zrakové pigmenty. fotorecepcia. Ktorá z mušlí očnej gule je spredu priehľadná. Bočné genikulárne telo je štruktúra
VIZUÁLNE PIGMENTY(lat. pigmentum paint) - fotosenzitívne pigmenty fotoreceptorov sietnice. Vnímaním energie svetelného impulzu 3. predmety prechádzajú zložitým fotochemickým cyklom. transformácie, v dôsledku ktorých sa samostatný zrakový receptor sietnice, obsahujúci 3. p.(kužeľ alebo tyčinka), dostáva do excitovaného stavu a prenáša prijatú informáciu pozdĺž zrakového nervu do c. n. od. Ako hlavná štrukturálna a funkčná časť fotoreceptorovej membrány zrakových buniek sietnice, 3. p. kľúčová úloha v mechanizmoch videnia (pozri).
Nomenklatúra a štruktúra zrakové pigmenty. Všetky študované 3. p. stavovcov a bezstavovcov sú komplexy vo vode nerozpustného membránového proteínu opsínu a s ním spojeného chromofóru (retinalu). Retinal alebo aldehyd vitamínu A môže existovať v dvoch formách, retinal1 a retinal2.
Podľa povahy chromofóru 3. je položka rozdelená do dvoch tried - rodopsíny (pozri), obsahujúce sietnicu1, a porfyropsíny obsahujúce sietnicu2. Rodopsíny sa nachádzajú v sietnici oka všetkých suchozemských a morských živočíchov, porfyropsíny sa nachádzajú v sietnici očí sladkovodných živočíchov. Niektoré ryby a obojživelníky majú v rovnakom čase 3 p.obsahujúce sietnicu! a sietnice. Existujú pokusy klasifikovať 3. p. na základe rozdielov v opsínoch špecifických pre tyčinky alebo čapíky sietnice. Napríklad rodopsín je komplex sietnice1 s tyčinkovým opsínom, jódpsín je komplex sietnice1 s čapičkovým opsínom, porfyropsín je sietnicový2 s tyčinkovým opsínom, komplex sietnica-čípkový opsín tvorí kyanopsín. Je však mimoriadne ťažké klasifikovať 3. položky na základe opsínov, keďže existuje najmenej päť rôznych opsínov.
Zo všetkých známych 3. položiek boli najkompletnejšie preskúmané rodopsíny izolované z očí býka, žaby a chobotnice. Ich prosba. hmotnosť (hmotnosť) rádovo 30-40 tisíc, každá molekula obsahuje cca. 400 aminokyselín a jeden chromofor. Okrem toho je v molekule 3. p. zahrnutý oligosacharidový reťazec: 3 glukozamínové radikály, 2 manóza, 1 galaktóza. Lipidy (ch. arr. fosfolipidy) tvoria silný komplex s molekulou 3. p. Pri zachovaní základných spektrálnych vlastností (pozri Spektrálna analýza) 3. položky bez lipidov strácajú množstvo funkčne dôležitých, napríklad schopnosť regenerácie.
Čistý retinal má žltú farbu, maximum jeho absorpčného spektra leží v oblasti 370 nm. Opsín je bezfarebný, absorpčné maximum je v ultrafialovej oblasti (cca 280 nm). Farba molekuly rodopsínu je červeno-ružová, maximálne absorpčné spektrum je cca. 500 nm. Dôvod takého silného spektrálneho posunu pri vzniku komplexu (od 370 do 500 nm – tzv. batochromický posun) zatiaľ nedostal jednoznačné vysvetlenie.
Maximá absorpčných spektier rodopsínov a porfyropsínov zachytávajú pomerne širokú oblasť viditeľného spektra - od 433 do 562 nm pre rodopsíny a od 510 do 543 nm pre porfyropsíny. Ak medzi porfyropsíny patria aj 3-p kužele žubrienka žaby, kapra a sladkovodnej korytnačky, teda cyanopsín s maximálnym absorpčným spektrom pri 620 nm, potom je táto oblasť ešte širšia. Rozvoj metód mikrospektrofotometrie umožnil stanoviť absorpčné spektrá mnohých typov jednotlivých fotoreceptorových buniek u zvierat a ľudí. Podľa získaných údajov majú 3. p. ľudskej sietnice nasledovné maximá absorpčného spektra: tyčinky 498, čapíky citlivé na modrú, zelenú a červenú - 440, 535 a 575 nm.
Štúdium 3. p. začalo v nemčine. výskumník H. Muller, ktorý v roku 1851 opísal, ako ružovo-fialová sietnica extrahovaná z oka žaby najprv na svetle zožltne a potom zbelie. V roku 1877 opísal tento jav aj F. Boll, ktorý dospel k záveru, že nejaký druh látky citlivej na červené svetlo sa nachádza vo zrakových bunkách sietnice a že zmena farby tejto látky je spojená s mechanizmom videnia. Veľkú zásluhu na štúdiu 3. p má Kuhne (W. Kuhne, 1877), Kromovi sa podarilo izolovať a podrobne preštudovať 3. p. Ním extrahovaný 3. p nazval vizuálnou fialovou, zistil jeho proteínovú povahu, skúmal niektoré jeho spektrálne vlastnosti a fototransformácie, objavil schopnosť 3 p. zotaviť sa v tme. K štúdiu 3. p. biochemik a fyziológ J. Wald.
Fototransformácie zrakových pigmentov. Vplyvom svetla na 3. p. v nich prebieha fotochemický cyklus. transformácie, ktoré sú založené na primárnej fotochemickej reakcii cis-trans izomerizácie sietnice (pozri Izoméria). V tomto prípade je väzba medzi chromoforom a proteínom narušená. Postupnosť premien 3. p. možno znázorniť takto: rodopsín (chromofor je v cis forme) -> prelumirhodopsín -> lumirhodopsín -> metarhodopsín I -> metarhodopsín II -> opsínový proteín -> chromofor v trans forme . Pod vplyvom enzýmu - retinoldehydrogenázy - tento prechádza na vitamín A, ktorý pochádza z vonkajších segmentov tyčiniek a čapíkov do buniek pigmentovej vrstvy sietnice. Pri zatemnení oka nastáva regenerácia 3. p., pre realizáciu rezu je potrebná prítomnosť cis-izoméru vitamínu A, ktorý slúži ako východiskový produkt pre tvorbu chromofóru (aldehyd vitamínu A). nevyhnutné. Pri nedostatku alebo absencii vitamínu A v organizme môže byť tvorba rodopsínu narušená a vzniká v dôsledku poruchy videnia za šera, tzv. nočná slepota (pozri Hemeralopia). V procese fototransformácie rodopsínu v štádiu prechodu lumirhodopsínu na metarhodopsín I v receptorovej bunke dochádza v reakcii na jasný záblesk tzv. skorý (krátkolatencia) receptorový potenciál. Zároveň nejde o vizuálny signál, hoci môže slúžiť ako jeden z testov na štúdium mechanizmu premien 3. p. v membráne fotoreceptora. Funkčnou hodnotou je tzv. neskorý receptorový potenciál, latentná perióda to-rogo (5-10 ms) je úmerná dobe tvorby metarhodopsínu II. Predpokladá sa, že reakcia prechodu metarhodopsínu I na metarodopsín II poskytuje výskyt vizuálny signál.
Keďže 3. predmety na svetle neustále menia farbu, musia existovať mechanizmy na ich neustále obnovenie. Niektoré z nich sú extrémne rýchle (fotoregenerácia), iné dosť rýchle (biochemické, regeneračné, tmavé), iné pomalé (syntéza 3. p. Pri neustálej obnove membrány fotoreceptora vo zrakovej bunke). Fotoregenerácia má fiziol, hodnotu u bezstavovcov (napr hlavonožce- chobotnica, chobotnica). V mechanizme biochem. regenerácia 3. p.u stavovcov hrá zrejme dôležitú úlohu enzým izomeráza (pozri), ktorý zabezpečuje izomerizáciu trans-retinalu (resp. trans-vitamínu A) opäť do cis-izomérnej formy. Neexistuje však žiadny definitívny dôkaz o existencii takéhoto enzýmu. Rovnaká reakcia tvorby molekuly 3. p. v prítomnosti 11-cis-izoméru sietnice a opsínu v systéme prebieha ľahko, bez výdaja energie. Zistila sa schopnosť bieleného rodopsínu k reakcii fosforylácie (pozri); predpokladá sa, že táto reakcia je jedným z článkov mechanizmu adaptácie zrakovej bunky na svetlo.
Bibliografia: Ackerman Yu Biofyzika, prekl. z angličtiny, M., 1964; Willie K. a Det e V. Biology, trans. z angličtiny, M., 1974, bibliografia; Konev S. V. a Volotovsky I. D. Úvod do molekulárnej fotobiológie, s. 61, Minsk, 1971; Ostrovsky M. A. a Fedorovich I. B. Fotoindukované zmeny fotoreceptorovej membrány, v knihe: Structure and functions of biol, membráns, ed. A. S. Troshina a ďalší, s. 224, M., 1975, bibliogr.; Fyziológia zmyslových systémov, vyd. G. V. Gershuni, časť 1, s. 88, L., 1971; Biochémia a fyziológia zrakových pigmentov, vyd. autor: H. Langer, B. a. o., 1973; Príručka senzorickej fyziológie, vyd. autor: H. A. R. Jung a. o., v. 7, pt 1-2, B., 1972.
M. A. Ostrovského.
vizuálny pigment štruktúrna a funkčná jednotka svetlocitlivej membrány fotoreceptorov (Pozri Fotoreceptory) sietnice - tyčinky a čapíky. V Z. p. sa uskutočňuje prvá etapa zrakového vnímania - absorpcia kvánt viditeľného svetla. Molekula Z. (molárna hmotnosť asi 40 000) pozostáva z chromofóru absorbujúceho svetlo a opsínu, komplexu bielkovín a fosfolipidov. Chromoforom všetkých Z. p. je aldehyd vitamínu A 1 alebo A 2 - retinal alebo 3-dehydroretinal. Dva typy opsínu (tyčinka a čapík) a dva typy sietnice, keď sa kombinujú v pároch, tvoria 4 typy z.p. nm), jódpsín (562 nm), porfyropsín (522 nm) a kyanopsín (620 nm). Primárna fotochemická väzba v mechanizme videnia (pozri Vision) spočíva vo fotoizomerizácii sietnice, ktorá pôsobením svetla mení svoju zakrivenú konfiguráciu na plochú. Po tejto reakcii nasleduje reťazec temných procesov vedúcich k objaveniu sa vizuálneho receptorového signálu, ktorý sa potom synapticky prenáša na ďalšie nervové elementy sietnice - bipolárne a horizontálne bunky. Lit.: Fyziológia zmyslových sústav, 1. časť, L., 1971, s. 88-125 (Manuál fyziológie); Wald G., Molekulárny základ vizuálnej excitácie, "Príroda", 1968, v. 219. M. A. Ostrovského.
Veľká sovietska encyklopédia. - M.: Sovietska encyklopédia. 1969-1978 .
Pozrite sa, čo je „vizuálny pigment“ v iných slovníkoch:
Konštrukčne funkčný. jednotka citlivá na svetlo. tyčinkové a kužeľové fotoreceptorové membrány v sietnici. Molekula 3. p. pozostáva z chromofóru absorbujúceho svetlo a opsínu z komplexu bielkovín a fosfolipidov. Chromofor je reprezentovaný aldehydom vitamínu A1 ... ... Biologický encyklopedický slovník
Rodopsín (vizuálna fialová) je hlavným vizuálnym pigmentom v tyčinkách ľudskej a zvieracej sietnice. Vzťahuje sa na komplexné proteíny, chromoproteíny. Modifikácie bielkovín charakteristické pre rôzne biologické druhy sa môžu výrazne líšiť ... Wikipedia
VIZUÁLNY (E) PIGMENT(Y)- pozri fotopigment... Slovník v psychológii
Pigment sietnice obsiahnutý vo vnútri tyčiniek, ktorý zahŕňa sietnicový (retinálny) vitamín A a proteín. Prítomnosť rodopsínu v sietnici je nevyhnutná pre normálne videnie v šere. Pod vplyvom svetla ... ... lekárske termíny
RHODOPSIN (RODOPSIN), FIALOVÝ VIZUÁL- (vizuálne fialový) retinálny pigment obsiahnutý vo vnútri tyčiniek, ktorý zahŕňa retinálny (retinálny) vitamín A a bielkoviny. Prítomnosť rodopsínu v sietnici je nevyhnutná pre normálne videnie v šere. Pod…… Výkladový slovník medicíny
- (vizuálna fialová), fotosenzitívna. komplexný proteín, zrakový pigment tyčinkových buniek v sietnici stavovcov a ľudí. Absorbovaním kvanta svetla (absorpčné maximum cca 500 nm) sa R. rozpadá a spôsobuje excitáciu ... ... Prírodná veda. encyklopedický slovník
- (zrakový pigment), svetlocitlivý tyčinkový proteín sietnice stavovcov a zrakových buniek bezstavovcov. R. glykoproteín (mol. m. cca 40 tis.; polypeptidový reťazec pozostáva z 348 aminokyselinových zvyškov), obsahujúci ... ... Chemická encyklopédia
- (z gr. ruže ródon a videnie ópsis) zraková fialová, hlavný vizuálny pigment tyčiniek sietnice stavovcov (okrem niektorých rýb a obojživelníkov v ranom štádiu vývoja) a bezstavovcov. Podľa chemikálií ...... Veľká sovietska encyklopédia
- (vizuálna fialová), komplexný proteín citlivý na svetlo, hlavný vizuálny pigment tyčinkových buniek sietnice u stavovcov a ľudí. Absorbovaním kvanta svetla (maximálna absorpcia je asi 500 nm) sa rodopsín rozkladá a spôsobuje ... ... encyklopedický slovník
Hlavný článok: Tyčinky (sietnica) Rodopsín (zastaraný, ale stále používaný názov vizuálna fialová) je hlavným vizuálnym pigmentom. Obsiahnuté v tyčinkách sietnice oka morských bezstavovcov, rýb, takmer všetkých suchozemských ... ... Wikipedia
25-10-2014, 13:04
Svetelné kvantá sú absorbované v receptoroch špecializovanými molekulami – vizuálnymi fotopigmentmi. Vizuálne pigmenty nezávisle na sebe objavili nemeckí fyziológovia F. Boll a W. Kuehne v rokoch 1877-1879. F. Boll si všimol, že sietnica izolovaná zo zorného skla žaby vyzerá najskôr ako červená a potom, keď na svetle bledne, zožltne a nakoniec je úplne bezfarebná.
W. Kuehne zistil, že v normálne fungujúcom oku zvieraťa sa farba sietnice po intenzívnom osvetlení svetlom obnoví, ak sa zviera opäť umiestni do tmy. Na základe týchto pozorovaní sa dospelo k záveru, že v sietnici oka sa nachádza fotosenzitívna látka (pigment), ktorej koncentrácia na svetle klesá a v tme sa obnovuje.
zrakové pigmenty- ide o komplexné molekuly chromolipoproteínov, ktoré sa u stavovcov a bezstavovcov skladajú z dvoch hlavných častí: chromofór (farbiaca časť molekuly, ktorá pri osvetlení určuje farbu receptora) a proteín (opsín). Chromofor určuje maximum a intenzitu absorpcie svetla v pigmente a je aldehydom alkoholov - vitamínov A1 a A2.
Názvy týchto aldehydov sú retinal-1 a retinal-2. Prítomnosť ďalšej dvojitej väzby v sietnici-2 vedie k posunu maxima absorpčného spektra do oblasti dlhých vlnových dĺžok. Retinal sa viaže na opsín, proteínovú mutáciu, ktorá sa nachádza iba vo fotoreceptoroch. Tyčinkový opsín, skotopsín a čapíky, fotopsín, sú izolované. Scotopsin pozostáva z 230-270 aminokyselinových zvyškov, ktorých sekvencia ešte nebola stanovená.
O fotopsíne je známe len málo: existujú iba údaje o jeho špirálovej konfigurácii. Skutočná časť molekuly pigmentu absorbujúca svetlo, sietnica, je typ karotenoidového pigmentu, najbežnejší vo fotosyntéze. tkanivá rastlín a živočíchov.
Spektrálne charakteristiky pigmentov sú určené kombináciami jedného zo sietníc s jedným alebo druhým druhom opsínov, ktorých rozmanitosť určuje rozmanitosť vizuálnych pigmentov u rôznych zvierat. Kombinácia skotopsínu s retinalom-1 teda vedie k vytvoreniu tyčinkového pigmentu - rodopsínu s absorpčným maximom pri? = 500 nm a jeho kombináciou s retinalom-2 vzniká ďalší tyčinkový pigment - porfyropsín s absorpčným maximom pri? = 525 nm. Kombinácie sietnice-1 s iný druh fotopsíny v sietnici ľudí a opíc tvoria kužeľové pigmenty s absorpčným maximom pri? = 445 nm, ? = 535 nm a? \u003d 570 nm, nazývané cyanolab, chlorolab a erythrolab.
Vlastnosti pigmentov absorbujúce svetlo sú presne určené interakciou iopsínového chromofóru: absorpčné maximá chromofóru a opsínu, brané oddelene, sú lokalizované v oblasti? \u003d 278 nm (retinal-1) a? = 378 nm (skotopsín), pričom ich zlúčenina - rodopsín - má absorpčné maximum pri? = 500 nm. Absorpčné maximum je jedným z parametrov spektrálnych charakteristík pigmentu, ktorý vo všeobecnosti určuje účinnosť absorpcie svetelných kvánt (fotónov) rôznych vlnových dĺžok pigmentom (obr. 3.1.4a).
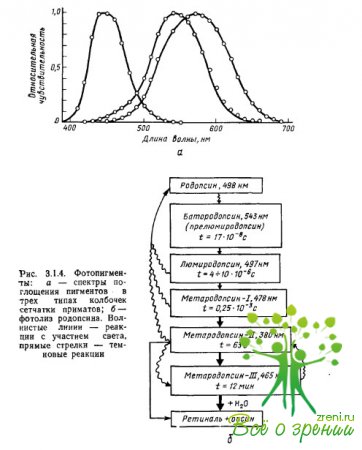
Prvé merania absorpčných spektier pigmentov v jednotlivých čapoch sietnice opíc a ľudí uskutočnili Brown, Wald a Marx a spol., s použitím metódy mikrospektrofotometrie špeciálne vyvinutej na tento účel. Prípravok sietnice bol umiestnený pod mikroskop, čo umožnilo pod vizuálnou kontrolou zamerať lúč svetla tej či onej vlnovej dĺžky na vonkajší segment fotoreceptora, ktorý obsahuje pigment.
Potom sa pre každú vlnovú dĺžku samostatne meralo množstvo svetla, ktoré prešlo kužeľom a nebolo ním absorbované (rozdiel medzi množstvom svetla dodaného do kužeľa a množstvom svetla ním prepusteného charakterizoval účinnosť absorpcie fotóny danej vlnovej dĺžky pigmentom). Merania sa opakovali po vyblednutí pigmentu v dôsledku intenzívneho osvetlenia (referenčné merania). Odpočítaním druhej série meraní od prvej sa získalo diferenciálne absorpčné spektrum pigmentu ako funkcia citlivosti na svetlo na vlnovej dĺžke žiarenia.
Z obr. 3.1.4a je možné vidieť, že pigmenty majú širokopásmové, silne sa prekrývajúce absorpčné spektrá. Všetky fotopigmenty majú dodatočnú absorpčnú charakteristiku karotenoidov (takzvaný nis-peak alebo "?-peak"), ktorá sa rovná 2/3 hlavného maxima. Pre pigmenty, ktorých hlavné absorpčné maximá sú lokalizované v častiach spektra s krátkymi a strednými vlnovými dĺžkami, vrchol cis spadá do ultrafialovej oblasti a neovplyvňuje analýzu žiarenia z viditeľného spektra. Ak sa ďalej pri konštrukcii funkcií spektrálnej citlivosti pigmentov stupnica vlnových dĺžok (abscisová os) prevedie na frekvenčnú stupnicu, potom sa absorpčné spektrá všetkých pigmentov, vrátane rôznych opsínov, ale rovnakého sieťového (napríklad sietnicového) -1), bude mať rovnaký a rovnaký tvar.
S touto prezentáciou údajov, ktorú prvýkrát navrhol Dartnall "(1953) pre pigmenty na báze sietnice-1, sa absorpčné spektrá rôznych fotopigmentov líšia iba v polohe maxima. A potom, keď poznáme absorpčné maximum pigmentu, môžeme obnoviť tvar jeho absorpčného spektra najskôr v súradniciach „frekvencia žiarenia - citlivosť“ a potom pomocou pomeru c = ?f a v súradniciach „vlnová dĺžka – citlivosť“.
Táto metóda obnovy absorpčného spektra neznámeho pigmentu z údajov iba o jeho maxime sa nazýva nomogramová metóda. Metóda nomogramov je tiež široko používaná v neurofyziológii farebného videnia pri riešení problému počtu a typu fotodetektorov konvergujúcich k jednému alebo druhému farebne citlivému neurónu: funkcia spektrálnej citlivosti neurónu sa porovnáva s jedným alebo iným nomogramom ( alebo ich kombinácia).
Rozmanitosť fotopigmentov v živočíšnej ríši koreluje s rozmanitosťou environmentálnych biotopov. U rýb a obojživelníkov môže sietnica obsahovať rodopsín aj porfyropsín, ktorých pomer sa počas života jedinca mení; u anadrómnych rýb predchádza prechodu zo slanej vody do sladkej vody nahradenie rodopsínu porfyropsínom a naopak. Spojenie zloženia pigmentu s ekológiou biotopu nie je rigidné: zmeny v zložení pigmentu môžu byť spôsobené sezónnymi zmenami a cirkadiánnymi rytmami.
Fotoizomerizácia.
Reakcia fotopigmentu na svetlo je určená počtom absorbovaných kvant, bez ohľadu na vlnovú dĺžku a energiu žiarenia: napríklad tie, ktoré absorbuje rodopsín. 10 kvant svetla pri? \u003d 430 nm v nej spôsobujú rovnaké štrukturálne zmeny (obr. 3.1.46), ako aj 10 absorbovaných kvantov svetla pri? = 500 nm.
A spektrálna selektivita (absorpčné spektrum) pigmentu je určená len rôznou pravdepodobnosťou absorpcie svetelných kvánt z rôznych častí viditeľného spektra. Zrakové pigmenty a v konečnom dôsledku aj fotoreceptory teda fungujú ako „počítače fotónov svetla“, ktoré sa navzájom líšia iba pravdepodobnosťou absorbovania fotónov rôznych vlnových dĺžok.
Keď jedna molekula rodopsínu pohltí 1 kvantum svetla, dôjde k zmene priestorovej konfigurácie jednej molekuly sietnice - fotoizomerizácii, ktorá spočíva v premene 11-cms formy sietnice na úplne premenu (obr. 3.1.4b). .
Fotoizomerizáciu sprevádza cyklus fotochemických reakcií, za ktorých dekódovanie bol G. Waldovi v roku 1930 udelená Nobelova cena. Konečnými produktmi fotoizomerizácie sú úplne traxretinal a opsín. Všetky medzireakcie až po krok metarodopsínu-II sú reverzibilné: produkty týchto reakcií sa môžu premeniť späť na rodopsín reverznou izomerizáciou gran-retinalu.
To znamená, že v molekule pigmentu pred metarodopsínom-II nenastávajú žiadne významné konformačné zmeny. V štádiu metarodopsínu-II sa sietnica oddelí od proteínovej časti molekuly. Zo všetkých etáp izomerizácie potrebuje svetlo iba prvá - premena rodopsínu na prelumirhodopsín (bathorhodopsín). Všetky ostatné reakcie prebiehajú v tme. Fotoizomerizácia prebieha podobne v kužeľových pigmentoch.
Fotoizomerizácia je teda molekulárny spúšťací (štartovací) mechanizmus fotorecepcie, ktorý spočíva v zmene priestorovej konfigurácie molekuly zrakového pigmentu. Ten objasňuje, prečo si príroda vybrala karotenoidy ako vizuálny pigment: prítomnosť dvojitých väzieb v sietnici určuje možnosť jeho izomerizácie a; jeho vysoká reaktivita. Okolo každej dvojitej väzby sa časti molekuly môžu otáčať a ohýbať v priestore. Táto forma je najmenej stabilná a najcitlivejšia na teplotu a svetlo. Nájdené v prírode rôzne druhy sietnica - 7, 9, 11, 13-cis-formy, 9,13-dicis-forma, ale vo zrakových pigmentoch je sietnica vždy v 11-cis-forme.
Resyntéza pigmentu vo fotoreceptoroch prebieha kontinuálne na svetle a v tme za účasti enzýmu retinénizomeráza a spočíva v reverznej izomerizácii retinolu z transformácie na 11-cis formu. V čapiciach sa tento enzým nachádza v samotných receptoroch, v tyčinkách, v pigmentovom epiteli, kam trans-retinol po oddelení od opsínu prechádza a kde sa izomerizuje na 11-cis formu. V tomto prípade sa vitamín A znovu použije a proteín sa syntetizuje v ribozómoch vnútorného segmentu.
Predpokladá sa, že k rekombinácii 11-cis-retinalu s opsínom dochádza spontánne. Pigmentový epitel obsahuje asi 11 % vitamínu A (retinolu) z 13 % v celom oku. Vitamín A vzniká v pečeni rozbitím reťazca karotenoidov obsiahnutých v potrave na dve časti a pridaním vody.
Vitamín A je transportovaný špeciálnym proteínom viažucim retinol do pigmentového epitelu, kde sa oxiduje na sietnicu. Pri nedostatku vitamínu A sa vyskytuje takzvaná „nočná slepota“ - zníženie absolútnej citlivosti na svetlo, ktoré ovplyvňuje najmä videnie za súmraku, prechádzajúce do trvalej slepoty v dôsledku deštrukcie proteínu, ktorý je vo voľnom stave nestabilný. Preto sa pri chorobe „nočnej slepoty“ odporúča jesť mrkvu s obsahom provitamínu A – betakaroténu.
Každý fotoreceptor obsahuje iba jeden fotopigment, ktorý sa vyznačuje tým či oným absorpčným spektrom. V tomto ohľade sú izolované kužele s krátkou, strednou a dlhou vlnovou dĺžkou, ktoré obsahujú pigmenty u primátov s absorpčnými maximami pri 445 nm, 535 nm a 570 nm. o sladkovodné ryby kužeľ s dlhou vlnovou dĺžkou obsahuje pigment s absorpčným maximom pri 620 nm, čo koreluje s tvarom spektrálneho rozloženia svetla v ich biotope.
Tieto čapíky sa niekedy označujú ako receptory citlivé na modrú, zelenú a červenú, podľa tradície siahajúcej až do trojzložkovej Young-Helmholtzovej hypotézy. Ale od druhej polovice 20. storočia, keď sa zistilo, že žiadny fotoreceptor sám o sebe nevykonáva spektrálnu analýzu, ale reaguje len na počet absorbovaných fotónov, bez ohľadu na vlnovú dĺžku žiarenia, sa tieto farebné názvy receptorov používajú iba v metaforickej podobe. zmysel.
Klasifikácia tyčiniek, napríklad u žaby, na "zelené" a "červené" v niektorých prípadoch nie je založená na spektrálnej citlivosti pigmentu v nich obsiahnutého, ale na farbe receptorových teliesok v prechádzajúcom svetle. „Zelené“ tyčinky teda obsahujú pigment s absorpčným maximom pri 432 nm, preto pri osvetlení bielym svetlom pohlcujú krátkovlnné žiarenie, prechádzajú všetkými ostatnými vlnovými dĺžkami a pri vizuálnej kontrole vyzerajú zelenkavo.
V sietnici rýb bol zaznamenaný vzťah medzi dĺžkou kužeľa a spektrálnym typom pigmentu, ktorý sa v ňom nachádza: najkratšie z nich obsahujú pigment s krátkou vlnovou dĺžkou a najdlhšie obsahujú pigment s dlhou vlnovou dĺžkou. Eberle naznačil, že by to mohol byť jeden z mechanizmov boja proti chromatickej aberácii. Ostrý obraz stredovlnného žiarenia je vďaka chromatickej aberácii umiestnený hlbšie (ďalej od šošovky) ako ostrý obraz krátkovlnného žiarenia a ohnisko pre dlhovlnné žiarenie je umiestnené ešte ďalej.
Stačí na príslušných úrovniach usporiadať krátkovlnné, strednovlnné a dlhovlnné receptory tak, aby všetky časti spektrálneho žiarenia boli rovnako ostro zamerané na receptory. To možno vysvetľuje skutočnosť, že v sietnici rýb jednotlivé čapíky s krátkymi segmentmi obsahujú pigment s krátkou vlnovou dĺžkou, zatiaľ čo v dvojitom kuželi najdlhší segment obsahuje pigment s dlhou vlnovou dĺžkou a kratší obsahuje pigment s krátkou vlnovou dĺžkou. alebo pigment strednej vlnovej dĺžky. Dvojité kužele so segmentmi rovnakej dĺžky obsahujú rovnaký pigment.
ωπς - oko) - spoločný názov viaceré zrakové pigmenty ľudí a niektorých cicavcov. Tieto pigmenty sa skladajú z proteínovej molekuly spojenej s chromoforom sietnice. Obsiahnuté v čapíkov sietnice a poskytujú farebné videnie, na rozdiel od iného zrakového pigmentu - rodopsínu, zodpovedného za videnie za šera.Zistilo sa, že zloženie vizuálnych pigmentov zahŕňa opsíny. Rôzne opsíny sa líšia svojim molekulárnym zložením aminokyselín a absorbujú svetlo pri mierne odlišných vlnových dĺžkach ako molekuly viazané na sietnicu.
Existenciu pigmentu kužeľa prvýkrát objavil (nepriamo) George Wald, ktorý mu dal meno. jodopsín. . V roku 1967 mu bola za tieto práce udelená Nobelova cena za fyziológiu a medicínu.
Dokonca aj Helmholtz veril, že v „terminálnom aparáte vlákien zrakového nervu“ (fotoreceptory sietnice) by mali existovať tri druhy fotochemicky rozložených látok, ktoré majú rôznu citlivosť vzhľadom na rôzne časti spektrum. .
Podľa trojzložkovej teórie videnia prevládajúcej v modernej vede sa verí, že existujú tri typy tohto pigmentu a sietnica obsahuje tri typy čapíkov, ktoré sú citlivé na modré, zelené a červené svetlo. V súlade s tým sa jodopsín, ktorý sa nachádza v čapiciach oka, skladá z troch pigmentov - chlorolabu, erytrolabu a kyanolabu; prvý z nich absorbuje lúče zodpovedajúce žltozelenej (absorpčné pásmo 450-630 nm), druhý - žlto-červený (500-700 nm) a tretí - modrozelený (500-700 nm) časti viditeľného svetla spektrum.
Typy vizuálnych pigmentov
Prvé pokusy nájsť tri pigmenty a podľa očakávania aj tri typy čapíkov (na základe predpokladov hypotézy trojzložkového videnia, že každý čapík obsahuje len jeden pigment) uskutočnil Rushton, ktorý zdokonalil denzitometrickú techniku pre in vivo. meranie absorpčných koeficientov svetla s rôznymi vlnovými dĺžkami - vo vrstve sietnicových fotoreceptorov. Ukázalo sa, že pri farebných abnormalitách chýba jeden z pigmentov prítomných u ľudí s normálnym zrakom: "erythrolab" (maximálne asi 585 nm.) v protanope a "chlorolab" (maximálne asi 540 nm.) v deuteranope.
Je potrebné poznamenať, že pri použití pojmu „prijímač“ sa v niektorých prípadoch celý súbor fotoreceptorov s rovnakou spektrálnou citlivosťou považuje za jeden prijímač; v iných prípadoch je otázka, či každý foveálny kužeľ obsahuje tri prijímač alebo len jeden. Zároveň nie je porušená prísnosť pojmu „prijímač“, ktorý v tomto prípade nezahŕňa špecifické morfologické znaky.
Ďalším krokom v tomto smere bolo štúdium fotopigmentov obsiahnutých v jednotlivých čapiciach človeka a makaka. Veľkosti foveálnych kužeľov sú príliš malé na to, aby mohli byť predmetom štúdia, a všetky získané údaje sa týkajú iba parafoveálnych kužeľov. Zdá sa, že každý čapík, aspoň ten extrafoveálny, obsahuje iba jeden z pigmentov, alebo prevažne jeden z nich.
Moderné metódy na štúdium fotopigmentov kužeľa
Denzitometria sietnicových buniek
→ denzitometria, môže byť nasledovná:
Ak prvé pokusy nájsť tri pigmenty a podľa očakávania aj tri typy kužeľov vykonal Rushton, ktorý túto techniku zdokonalil Denzitometria na meranie absorpčných koeficientov svetla s rôznymi vlnovými dĺžkami - vo vrstve sietnicových fotoreceptorov a ukázalo sa, že pri farebných abnormalitách chýba jeden z pigmentov, ktoré majú ľudia s normálnym zrakom: "erythrolab" (maximálne asi 585 nm.) v protanope a " chlorolab" (maximálne asi 540 nm.) - v deuteranop, teraz pomocou špeciálnych metód dystometrie pomocou moderných denzitometrov, vedci dokážu určiť prácu kužeľov a tyčí v stave normálnej činnosti a diagnostikovať ich choroby.
pozri tiež
Zdroje
Literatúra
- Khokhlova T.V. Moderné predstavy o vízii cicavcov // Journal of General Biology. Ročník 73, 2012. Číslo 6, november-december. Stránka 418-434.


