Pigments visuels. photoréception. Laquelle des membranes du globe oculaire est transparente devant. Le corps géniculé latéral est une structure
PIGMENTS VISUELS(lat. pigmentum paint) - pigments photosensibles des photorécepteurs de la rétine. En percevant l'énergie d'une impulsion lumineuse, 3. les objets subissent un cycle photochimique complexe. transformations, à la suite desquelles un récepteur visuel distinct de la rétine, contenant 3. p. (cône ou bâtonnet), entre dans un état excité et transmet les informations reçues au système nerveux central le long du nerf optique. n. Avec. Étant la principale partie structurelle et fonctionnelle de la membrane photoréceptrice des cellules visuelles de la rétine, 3. p. jouent ainsi rôle clé dans les mécanismes de la vision (voir).
Nomenclature et structure pigments visuels. Tous les 3.p étudiés chez les animaux vertébrés et invertébrés sont des complexes de la protéine membranaire insoluble dans l'eau, l'opsine, et du chromophore associé (rétinien). Le rétinal, ou aldéhyde de vitamine A, peut exister sous deux formes : rétinal1 et rétinal2.
Selon la nature du chromophore, les 3. p. sont divisés en deux classes - les rhodopsines (voir), contenant du rétinal1, et les porphyropsines, contenant du rétinal2. Les rhodopsines se trouvent dans la rétine des yeux de tous les animaux terrestres et marins, les porphyropsines se trouvent dans la rétine des yeux des animaux d'eau douce. Chez certains poissons et amphibiens, 3. éléments contenant simultanément du rétinien ! et rétinienne. Il existe des tentatives pour classer 3.p. sur la base de différences d'opsines spécifiques aux bâtonnets ou cônes de la rétine. Par exemple, la rhodopsine est un complexe de rétinal1 avec l'opsine en bâtonnet, l'iodopsine est un complexe de rétinal1 avec l'opsine en cône, la porphyropsine est un complexe rétinien2 avec l'opsine en bâtonnet, le complexe opsine rétinien-cône forme la cyanopsine. Cependant, il est extrêmement difficile de classer 3.p. sur la base des opsines, puisqu'il existe au moins cinq opsines différentes.
De toutes les 3. p. connues, ce sont les rhodopsines isolées des yeux de taureaux, de grenouilles et de calmars qui ont été les plus étudiées. Ils disent le poids (masse) est d'environ 30 à 40 000, chaque molécule contient env. 400 acides aminés et un chromophore. De plus, la molécule 3.p. comprend une chaîne oligosaccharide : 3 radicaux glucosamine, 2 mannose, 1 galactose. Les lipides (principalement les phospholipides) forment un complexe fort avec la molécule 3.p. Tout en conservant leurs propriétés spectrales de base (voir Analyse spectrale), les 3.p. sans lipides perdent un certain nombre de propriétés fonctionnellement importantes, par exemple la capacité de récupération.
Le rétinien pur a une couleur jaune, le maximum de son spectre d'absorption se situe aux alentours de 370 nm. L'opsine est incolore, l'absorption maximale se situe dans la région ultraviolette (environ 280 nm). La couleur de la molécule de rhodopsine est rose rougeâtre, le spectre d'absorption maximal est d'env. 500 nm. La raison d'un déplacement spectral aussi fort lors de la formation du complexe (de 370 à 500 nm - ce qu'on appelle le déplacement bathochrome) n'a pas encore reçu d'explication sans ambiguïté.
Les spectres d'absorption maxima des rhodopsines et des porphyropsines couvrent une région assez large du spectre visible - de 433 à 562 nm pour les rhodopsines et de 510 à 543 nm pour les porphyropsines. Si les 3èmes cônes du têtard de la grenouille, de la carpe et de la tortue d'eau douce sont également considérés comme des porphyropsines, c'est-à-dire des cyanopsines avec un spectre d'absorption maximal à 620 nm, alors cette région s'avère encore plus large. Le développement des méthodes de microspectrophotométrie a permis de déterminer les spectres d'absorption de nombreux types de cellules photoréceptrices uniques chez l'animal et l'homme. Selon les données obtenues, les 3.p. de la rétine humaine ont les spectres d'absorption maxima suivants : bâtonnets 498, cônes sensibles au bleu, au vert et au rouge - 440, 535 et 575 nm, respectivement.
L'étude de 3. P. a commencé en allemand. le chercheur H. Muller, qui a décrit en 1851 comment la rétine violet rosé, extraite de l'œil d'une grenouille, devient jaunâtre puis blanchâtre à la lumière. En 1877, F. Boll a également décrit ce phénomène, concluant que dans les cellules visuelles de la rétine se trouve une sorte de substance sensible à la lumière rouge et que la décoloration de cette substance est associée au mécanisme de la vision. Une grande partie du mérite de l'étude des 3e points revient à Kuhne (W. Kuhne, 1877), qui a réussi à isoler les 3e points et à les étudier en détail. Il a nommé le 3.p. dont il a extrait le violet visuel, a établi sa nature protéique, a étudié certaines de ses propriétés spectrales et ses phototransformations, et a découvert la capacité du 3.p. à être restauré dans l'obscurité. Amer. a apporté une grande contribution à l'étude de 3. p. biochimiste et physiologiste J. Wald.
Phototransformations des pigments visuels. Lorsque 3.p. est exposé à la lumière, un cycle photochimique s’y produit. transformations, qui sont basées sur la réaction photochimique primaire d'isomérisation cis-trans du rétinien (voir Isomérie). Dans ce cas, la connexion entre le chromophore et la protéine est perturbée. La séquence de transformations de 3. p. peut se présenter comme suit : rhodopsine (le chromophore est sous forme cis) -> prélumirodopsine -> lumyrhodopsine -> métarhodopsine I -> métarhodopsine II -> protéine opsine -> chromophore sous forme trans. Sous l'influence de l'enzyme - la rétinol déshydrogénase - cette dernière est transformée en vitamine A, qui provient des segments externes des bâtonnets et des cônes et pénètre dans les cellules de la couche pigmentaire de la rétine. Lorsque l'œil s'assombrit, la régénération du 3.p. se produit ; pour réaliser la coupe, la présence de l'isomère cis de la vitamine A est nécessaire, qui sert de produit de départ pour la formation du chromophore (aldéhyde de vitamine A). En cas de carence ou d'absence de vitamine A dans le corps, la formation de rhodopsine peut être perturbée et, par conséquent, un trouble de la vision crépusculaire, ce qu'on appelle, peut se développer. cécité nocturne (voir Héméralopie). Dans le processus de phototransformations de la rhodopsine au stade de la transition de la lumirhodopsine à la métarhodopsine I dans la cellule réceptrice se produit en réponse à un éclair lumineux de ce qu'on appelle. potentiel de récepteur précoce (latence courte). En même temps, ce n'est pas un signal visuel, bien qu'il puisse servir d'un des tests pour étudier le mécanisme des transformations 3.p. dans la membrane photoréceptrice. La signification fonctionnelle est ce qu'on appelle. potentiel de récepteur tardif, dont la période de latence (5-10 ms) est comparable au temps de formation de la métarhodopsine II. On suppose que la réaction de transition de la métarhodopsine I en métarhodopsine II permet l'émergence signal visuel.
Puisque les 3.p. sont continuellement décolorées à la lumière, il doit y avoir des mécanismes pour leur restauration constante. Certains d'entre eux sont extrêmement rapides (photorégénération), d'autres sont assez rapides (biochimiques, régénération, Dark), d'autres sont lents (synthèse de 3. p. lors du renouvellement constant de la membrane photoréceptrice de la cellule visuelle). La photorégénération est fiziol, importante chez les animaux invertébrés (par exemple, céphalopodes- calamar, poulpe). Dans le mécanisme biochimique Dans la régénération de 3. p. chez les vertébrés, l'enzyme isomérase (voir), qui assure l'isomérisation du trans-rétinien (ou trans-vitamine A) à nouveau sous la forme cis-isomère, joue apparemment un rôle important. Cependant, aucune preuve définitive de l’existence d’une telle enzyme n’est encore disponible. La réaction même de formation de la molécule 3.p. en présence de l'isomère 11-cis du rétinal et de l'opsine se produit facilement, sans consommation d'énergie. La capacité de la rhodopsine blanchie à subir une réaction de phosphorylation a été découverte (voir) ; on suppose que cette réaction est l'un des maillons du mécanisme d'adaptation lumineuse de la cellule visuelle.
Bibliographie: Akkerman Yu. Biophysique, trad. de l'anglais, M., 1964 ; Willie K. et Det e V. Biologie, trans. de l'anglais, M., 1974, bibliogr.; Konev S.V. et Volotovsky I.D. Introduction à la photobiologie moléculaire, p. 61, Minsk, 1971 ; Ostrovsky M. A. et Fedorovich I. B. Modifications photo-induites dans la membrane photoréceptrice, dans le livre : Structure et fonctions du biol, membranes, éd. A.S. Troshina et al., p. 224, M., 1975, bibliogr.; Physiologie des systèmes sensoriels, éd. G.V. Gershuni, partie 1, p. 88, L., 1971 ; Biochimie et physiologie des pigments visuels, éd. par H. Langer, V. a. o., 1973 ; Manuel de physiologie sensorielle, éd. par H.A.R. Jung a. o., v. 7, parties 1-2, B., 1972.
M.A. Ostrovsky.
Pigment visuel unité structurelle et fonctionnelle de la membrane photosensible des photorécepteurs (Voir Photorécepteurs) de la rétine - bâtonnets et cônes. Dans le champ visuel, se produit la première étape de la perception visuelle : l’absorption des quanta de lumière visible. La molécule Zp (masse molaire environ 40 000) est constituée d'un chromophore qui absorbe la lumière et de l'opsine, un complexe de protéines et de phospholipides. Le chromophore de tous les minéraux est l'aldéhyde de la vitamine A 1 ou A 2 - rétinienne ou 3-déhydrorétinienne. Deux types d'opsine (bâtonnet et cône) et deux types de rétiniens, lorsqu'ils sont combinés par paires, forment 4 types d'opsine, différant par leur spectre d'absorption : la rhodopsine (la protéine en bâtonnet la plus courante) ou le violet visuel (absorption maximale 500 nm), l'iodopsine (562 nm), la porphyropsine (522 nm) et la cyanopsine (620 nm). Le principal lien photochimique du mécanisme de la vision (voir Vision) consiste en la photoisomérisation de la rétine qui, sous l'influence de la lumière, change sa configuration incurvée en une configuration plate. Cette réaction est suivie d'une chaîne de processus sombres conduisant à l'apparition d'un signal de récepteur visuel, qui est ensuite transmis synaptiquement aux éléments neuronaux suivants de la rétine - les cellules bipolaires et horizontales. Lit. : Physiologie des systèmes sensoriels, partie 1, L., 1971, p. 88-125 (Manuel de physiologie) ; Wald G., Les bases moléculaires de l'excitation visuelle, « Nature », 1968, v. 219. M.A. Ostrovsky.
Grande Encyclopédie soviétique. - M. : Encyclopédie soviétique. 1969-1978 .
Voyez ce qu'est « pigment visuel » dans d'autres dictionnaires :
Structurel et fonctionnel L'appareil est sensible à la lumière. membranes des photorécepteurs rétiniens des bâtonnets et des cônes. La molécule 3.p est constituée d'un chromophore absorbant la lumière et d'un complexe opsine de protéines et de phospholipides. Le chromophore est représenté par l'aldéhyde de la vitamine A1... ... Dictionnaire encyclopédique biologique
La rhodopsine (violet visuel) est le principal pigment visuel des bâtonnets de la rétine des humains et des animaux. Fait référence à des protéines complexes - les chromoprotéines. Les modifications protéiques caractéristiques de différentes espèces biologiques peuvent varier considérablement... Wikipédia
PIGMENT(S) VISUEL(S)- Voir photopigment... Dictionnaire en psychologie
À l’intérieur des bâtonnets se trouve le pigment rétinien de l’œil, qui comprend du rétinaldéhyde (rétinien), de la vitamine A et des protéines. La présence de rhodopsine dans la rétine est nécessaire à une vision normale dans des conditions de faible luminosité. Sous l'influence de la lumière... ... Termes médicaux
RHODOPSINE, VISUEL VIOLET- (violet visuel) pigment rétinien contenu à l'intérieur des bâtonnets, qui comprend du rétinaldéhyde (rétinien), de la vitamine A et des protéines. La présence de rhodopsine dans la rétine est nécessaire à une vision normale dans des conditions de faible luminosité. Sous… … Dictionnaire explicatif de la médecine
- (violet visuel), sensible à la lumière. protéine complexe, basique pigment visuel des cellules en bâtonnets de la rétine chez les vertébrés et les humains. En absorbant un quantum de lumière (absorption maximale environ 500 nm), R. se désintègre et provoque une excitation... ... Sciences naturelles. Dictionnaire encyclopédique
- (pigment visuel), une protéine photosensible des bâtonnets de la rétine des vertébrés et des cellules visuelles des invertébrés. R. glycoprotéine (poids mol. environ 40 000 ; la chaîne polypeptidique est constituée de 348 résidus d'acides aminés), contenant... ... Encyclopédie chimique
- (du grec rhódon rose et opsis vision) violet visuel, principal pigment visuel des bâtonnets de la rétine des vertébrés (à l'exception de certains poissons et amphibiens aux premiers stades de développement) et des animaux invertébrés. Selon le produit chimique... ... Grande Encyclopédie Soviétique
- (violet visuel), une protéine complexe photosensible, principal pigment visuel des bâtonnets de la rétine chez les vertébrés et l'homme. En absorbant un quantum de lumière (absorption maximale environ 500 nm), la rhodopsine se désintègre et provoque... ... Dictionnaire encyclopédique
Article principal : Bâtonnets (rétine) La rhodopsine (un nom obsolète mais toujours utilisé pour le violet visuel) est le principal pigment visuel. Contenu dans les bâtonnets rétiniens des invertébrés marins, des poissons, presque tous terrestres... ... Wikipédia
25-10-2014, 13:04
Les quanta de lumière sont absorbés dans les récepteurs par des molécules spécialisées : les photopigments visuels. Les pigments visuels ont été découverts indépendamment par les physiologistes allemands F. Boll et W. Kühne en 1877-1879. F. Boll a remarqué que la rétine isolée de la cupule optique d'une grenouille paraît d'abord rouge, puis, s'estompant à la lumière, devient jaune et enfin complètement incolore.
W. Kuehne a découvert que dans l'œil d'un animal fonctionnant normalement, la couleur de la rétine après un éclairage intense par la lumière est restaurée si l'animal est à nouveau placé dans l'obscurité. Sur la base de ces observations, il a été conclu qu'il existe une substance photosensible (pigment) dans la rétine de l'œil, dont la concentration diminue à la lumière et se rétablit dans l'obscurité.
Pigments visuels- ce sont des molécules complexes de chromolipoprotéines, constituées de deux parties principales chez les vertébrés et les invertébrés : un chromophore (la partie colorante de la molécule qui détermine la couleur du récepteur lorsqu'il est éclairé) et une protéine (opsine). Le chromophore détermine le maximum et l'intensité de l'absorption de la lumière dans le pigment et est un aldéhyde d'alcools - vitamines A1 et A2.
Les noms de ces aldéhydes sont rétinal-1 et rétinal-2. La présence d'une double liaison supplémentaire dans le rétinal-2 entraîne un déplacement du spectre d'absorption maximal vers des longueurs d'onde plus longues. La rétine s'attache à l'opsine, une mutation protéique présente uniquement dans les photorécepteurs. L'opsine en bâtonnet est la scotopsine et l'opsine en cône est la photopsine. La scotopsine est constituée de 230 à 270 résidus d'acides aminés dont la séquence n'a pas encore été déterminée.
On sait peu de choses sur la photopsine : il existe uniquement des données sur sa configuration hélicoïdale. La partie réelle de la molécule de pigment qui absorbe la lumière, la rétine, est un type de pigment caroténoïde, le plus courant dans la photosynthèse. tissus de plantes et d'animaux.
Les caractéristiques spectrales des pigments sont déterminées par des combinaisons de l'un des rétiniens avec l'un ou l'autre type d'opsine, dont la diversité détermine la diversité des pigments visuels chez différents animaux. Ainsi, la combinaison de la scotopsine avec le rétinal-1 conduit à la formation d'un pigment en bâtonnets - la rhodopsine avec un maximum d'absorption à ? = 500 nm, et sa combinaison avec le rétinal-2 forme un autre pigment en bâtonnets - la porphyropsine avec un maximum d'absorption à ? =525nm. Combinaisons de rétinien-1 avec différents types les photopsines dans la rétine des humains et des singes forment des pigments coniques avec des maxima d'absorption à ? = 445 nm, ? = 535 nm et ? =570 nm, appelés respectivement cyanolab, chlorolab et erythrolab.
Les propriétés d'absorption de la lumière des pigments sont déterminées précisément par l'interaction du chromophore iopsine : les maxima d'absorption du chromophore et de l'opsine, pris séparément, sont localisés dans la région ? =278 nm (rétinien-1) et ? = 378 nm (scotopsine), alors que leur composé - la rhodopsine - a un maximum d'absorption à ? = 500 nm. L'absorption maximale est l'un des paramètres des caractéristiques spectrales d'un pigment, qui détermine généralement l'efficacité d'absorption des quanta de lumière (photons) par le pigment. différentes longueurs vagues (Fig. 3.1.4a).
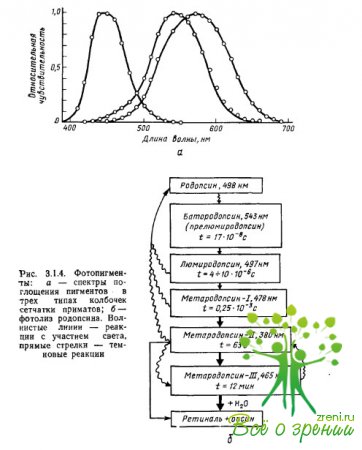
Les premières mesures des spectres d'absorption des pigments dans les cônes individuels de la rétine du singe et de l'humain ont été réalisées par Brown, Wald et Marx et al., en utilisant une méthode de microspectrophotométrie spécialement développée à cet effet. La préparation rétinienne a été placée sous un microscope, ce qui a permis, sous contrôle visuel, de focaliser un faisceau lumineux d'une longueur d'onde particulière sur le segment externe du photorécepteur, où est contenu le pigment.
Ensuite, pour chaque longueur d'onde, des mesures ont été prises séparément de la quantité de lumière qui traversait le cône et n'était pas absorbée par celui-ci (la différence entre la quantité de lumière fournie au cône et la quantité de lumière transmise par celui-ci caractérisait l'efficacité de l'absorption par le pigment des photons d'une longueur d'onde donnée). Les mesures ont été répétées après la décoloration du pigment due à un éclairage intense (mesures de référence). En soustrayant la deuxième série de mesures de la première, un spectre d'absorption différentielle du pigment a été obtenu en fonction de la sensibilité à la lumière sur la longueur d'onde du rayonnement.
De la fig. 3.1.4a, il est clair que les pigments ont des spectres d’absorption à large bande et fortement chevauchants. Tous les photopigments ont un maximum d'absorption supplémentaire caractéristique des caroténoïdes (appelé pic de schis ou ?-pic), égal aux 2/3 du maximum principal. Pour les pigments dont les principaux maxima d'absorption sont localisés dans les parties du spectre à longueur d'onde courte et moyenne, le pic cis tombe dans la région ultraviolette et n'affecte pas l'analyse du rayonnement du spectre visible. Si, en outre, lors de la construction des fonctions de sensibilité spectrale des pigments, l'échelle de longueur d'onde (axe des x) est convertie en échelle de fréquence, alors les spectres d'absorption de tous les pigments, y compris les différentes opsines, mais le même rétial (par exemple, rétinien-1), aura la même forme.
Avec cette représentation des données, proposée pour la première fois par Dartnall (1953) pour les pigments à base de rétinal-1, les spectres d'absorption des différents photopigments ne diffèrent les uns des autres que par la position du maximum. Et puis, connaissant l'absorption maximale du pigment, on peut restituer la forme de son spectre d'absorption d'abord dans les coordonnées « fréquence de rayonnement - sensibilité », puis, en utilisant le rapport c = ?f et dans les coordonnées « longueur d'onde - sensibilité ».
Cette méthode de reconstruction du spectre d'absorption d'un pigment inconnu à partir de données uniquement sur son maximum est appelée méthode du nomogramme. La méthode du nomogramme est également largement utilisée en neurophysiologie de la vision des couleurs pour décider du nombre et du type de photodétecteurs qui convergent vers un neurone sensible à la couleur particulier : la fonction de sensibilité spectrale d'un neurone est comparée à l'un ou l'autre nomogramme (ou une combinaison celui-ci).
La diversité des photopigments dans le monde animal est en corrélation avec la diversité des conditions environnementales. Chez les poissons et les amphibiens, la rétine peut contenir à la fois de la rhodopsine et de la porphyropsine, dont le rapport change au cours de la vie de l'individu ; Chez les poissons anadromes, le remplacement de la rhodopsine par la porphyropsine précède le passage de l'eau salée à l'eau douce et vice versa. Le lien entre l'ensemble des pigments et l'écologie de l'habitat n'est pas strict : des changements dans la composition des pigments peuvent être provoqués par des changements saisonniers et des rythmes circadiens.
Photoisomérisation.
La réaction d'un photopigment à la lumière est déterminée par le nombre de quanta absorbés, quelles que soient la longueur d'onde et l'énergie du rayonnement : par exemple ceux absorbés par la rhodopsine. 10 quanta de lumière à ? =430 nm provoquent les mêmes changements structurels (Fig. 3.1.46) que 10 lumière quantique absorbée à ? = 500 nm.
Et la sélectivité spectrale (spectre d'absorption) du pigment est déterminée uniquement par la probabilité différente d'absorption des quanta de lumière provenant de différentes parties du spectre visible. Ainsi, les pigments visuels et, en fin de compte, les photorécepteurs fonctionnent comme des « compteurs de quanta de lumière », ne différant que par la probabilité d’absorber des photons de différentes longueurs d’onde.
Lorsqu'une molécule de rhodopsine absorbe 1 quantum de lumière, un changement dans la configuration spatiale d'une molécule rétinienne se produit - la photoisomérisation, qui consiste en la transformation de la forme rétinienne de 11 cm en une forme complètement transformée (Fig. 3.1.4b).
La photoisomérisation s'accompagne d'un cycle de réactions photochimiques, pour le déchiffrement desquelles G. Wald reçut le prix Nobel en 1930. Les produits finaux de la photoisomérisation sont le trexrétinal et l'opsine. Toutes les réactions intermédiaires jusqu'au stade métarhodopsine-P sont réversibles : les produits de ces réactions peuvent être reconvertis en rhodopsine par isomérisation gran-rétinienne inverse.
Cela signifie qu’avant la métarhodopsine-II, aucun changement conformationnel significatif ne se produit dans la molécule pigmentaire. Au stade de la métarhodopsine-II, le rétinien se détache de la partie protéique de la molécule. De toutes les étapes de l'isomérisation, seule la première - la transformation de la rhodopsine en prélumirhodopsine (batorhodopsine) - nécessite de la lumière. Toutes les autres réactions ont lieu dans l’obscurité. La photoisomérisation se produit de la même manière dans les pigments coniques.
Ainsi, la photoisomérisation est un mécanisme moléculaire déclencheur de la photoréception, consistant en un changement dans la configuration spatiale de la molécule du pigment visuel. Ce dernier explique pourquoi la nature a choisi les caroténoïdes comme pigment visuel : la présence de doubles liaisons dans la rétine détermine la possibilité de son isomérisation et ; sa grande réactivité. Autour de chaque double liaison, des parties de la molécule peuvent tourner et se plier dans l’espace. Cette forme est la moins stable et la plus sensible à la température et à la lumière. Trouvé dans la nature différentes sortes rétinien - formes 7, 9, 11, 13-cis, forme 9,13-dicis, mais dans les pigments visuels, la rétine est toujours sous la forme 11-cis.
La resynthèse pigmentaire dans les photorécepteurs se produit en continu à la lumière et dans l'obscurité avec la participation de l'enzyme rétinène isomérase et consiste en l'isomérisation inverse du rétinol de la transformation à la forme 11-cis. Dans les cônes, cette enzyme est localisée dans les récepteurs eux-mêmes, dans les bâtonnets, dans l'épithélium pigmentaire, où, après séparation de l'opsine, passe le trans-rétinol et où s'effectue son isomérisation sous la forme 11-cis. Dans ce cas, la vitamine A rétinienne est réutilisée et la protéine est synthétisée dans les ribosomes du segment interne.
On suppose que la recombinaison du 11-cis-rétinien avec l'opsine se produit spontanément. Environ 11 % de la vitamine A (rétinol) est localisée dans l’épithélium pigmentaire, sur 13 % dans l’œil entier. La vitamine A est formée dans le foie en brisant la chaîne caroténoïde présente dans les aliments en deux parties et en ajoutant de l'eau.
La vitamine A est transportée par une protéine spéciale liant le rétinol jusqu'à l'épithélium pigmentaire, où elle est oxydée en rétinien. En cas de carence en vitamine A, il se produit ce qu'on appelle la « cécité nocturne » - une diminution de la sensibilité absolue à la lumière, affectant particulièrement la vision au crépuscule, se transformant en cécité permanente en raison de la destruction d'une protéine instable à l'état libre. Par conséquent, si vous souffrez de cécité nocturne, il est recommandé de manger des carottes contenant de la provitamine A – bêta-carotène.
Chaque photorécepteur ne contient qu'un seul photopigment, caractérisé par un spectre d'absorption particulier. À cet égard, on distingue les cônes à ondes courtes, moyennes et longues, qui chez les primates contiennent des pigments avec des maxima d'absorption à 445 nm, 535 nm et 570 nm, respectivement. U poisson d'eau douce Le cône à ondes longues contient un pigment avec un maximum d'absorption à 620 nm, qui est en corrélation avec la forme de la distribution spectrale de la lumière dans leur habitat.
Parfois, selon la tradition héritée de l'hypothèse des trois composantes de Young-Helmholtz, ces cônes sont appelés récepteurs sensibles au bleu, au vert et au rouge. Mais depuis la seconde moitié du XXe siècle, lorsqu'il a été constaté qu'aucun photorécepteur lui-même n'effectue une analyse spectrale, mais réagit uniquement au nombre de photons absorbés, quelle que soit la longueur d'onde du rayonnement, ces noms de couleur des récepteurs ne sont utilisés que dans un sens métaphorique.
La classification des bâtonnets, par exemple chez une grenouille, en « vert » et « rouge » dans certains cas est basée non pas sur la sensibilité spectrale du pigment qu'ils contiennent, mais sur la couleur des corps récepteurs en lumière transmise. Ainsi, les bâtonnets « verts » contiennent un pigment avec une absorption maximale à 432 nm. Par conséquent, lorsqu'ils sont éclairés par de la lumière blanche, ils absorbent le rayonnement à ondes courtes, transmettant toutes les autres longueurs d'onde, et lors d'une inspection visuelle, ils apparaissent verdâtres.
Dans la rétine des poissons, une relation a été notée entre la longueur du cône et le type spectral de pigment qu'il contient : les plus courts d'entre eux contiennent des pigments de courte longueur d'onde, et les plus longs, de longue longueur d'onde. Eberly a suggéré que cela pourrait être l'un des mécanismes permettant de lutter contre l'aberration chromatique. En raison de l'aberration chromatique, l'image nette du rayonnement à ondes moyennes est située plus profondément (plus loin de l'objectif) que l'image nette du rayonnement à ondes courtes, et le foyer du rayonnement à ondes longues est localisé encore plus loin.
Il suffit de placer les récepteurs à ondes courtes, moyennes et longues à des niveaux appropriés pour que toutes les parties du rayonnement spectral soient focalisées avec la même netteté sur les récepteurs. Cela peut expliquer le fait que dans la rétine des poissons, les cônes simples avec des segments courts contiennent des pigments de courte longueur d'onde, et que dans le double cône, le segment le plus long contient un pigment de longue longueur d'onde et le plus court contient un pigment de courte ou moyenne longueur d'onde. . Les doubles cônes avec des segments de même longueur contiennent le même pigment.
ωπς - œil) - Nom commun plusieurs pigments visuels des humains et de certains mammifères. Ces pigments sont constitués d'une molécule protéique liée au chromophore rétinien. Contenu dans les cônes de la rétine et assure la vision des couleurs, contrairement à un autre pigment visuel - la rhodopsine, responsable de la vision crépusculaire.Il a été établi que les pigments visuels comprennent les opsines. Différentes opsines diffèrent par les acides aminés de leurs molécules et absorbent la lumière à des longueurs d'onde légèrement différentes, comme les molécules liées à la rétine.
L'existence du pigment conique a été découverte pour la première fois (indirectement) par George Wald, qui lui a donné son nom. iodopsine. . En 1967, il reçut le prix Nobel de physiologie ou médecine pour ces travaux.
Helmholtz pensait également que dans « l'appareil terminal des fibres du nerf optique » (photorécepteurs de la rétine), il devrait y avoir trois types de substances en décomposition photochimique qui ont une sensibilité différente à Différents composants spectre .
Selon la théorie de la vision à trois composants qui prévaut dans la science moderne, on pense qu'il existe trois types de ce pigment et que la rétine contient donc trois types de cônes sensibles à la lumière bleue, verte et rouge. En conséquence, l'iodopsine, située dans les cônes de l'œil, est constituée de trois pigments : le chlorolab, l'érythrolab et le cyanolab ; le premier d'entre eux absorbe les rayons correspondant au jaune-vert (bande d'absorption 450-630 nm), le deuxième - jaune-rouge (500-700 nm) et le troisième - bleu-vert (500-700 nm) partie du visible spectre.
Types de pigments visuels
Les premières tentatives pour trouver trois pigments et, comme prévu, trois types de cônes (basées sur l'hypothèse de la vision à trois composants selon laquelle chaque cône ne contient qu'un seul pigment) ont été réalisées par Rushton, qui a perfectionné la technique de densitométrie pour la mesure intravitale. des coefficients d'absorption de la lumière avec différentes longueurs d'onde - dans la couche photorécepteurs rétiniens. Il a été démontré que les anomalies de couleur sont dépourvues d'un des pigments présents chez les personnes ayant une vision normale : « érythrolab » (maximum environ 585 nm) dans le protanope et « chlorolab » (maximum environ 540 nm) dans le deutéranope.
Il convient de noter que lorsqu'on utilise le terme «récepteur», dans certains cas, l'ensemble des photorécepteurs ayant la même sensibilité spectrale est considéré comme un seul. destinataire; dans d'autres cas, la question de savoir si chaque cône fovéal contient trois destinataire ou juste un. Dans le même temps, la rigueur du concept de «récepteur» n'est pas violée, qui dans ce cas n'inclut pas de caractéristiques morphologiques spécifiques.
L'étape suivante dans cette direction a été l'étude des photopigments contenus dans les cônes individuels des humains et des macaques. Les cônes fovéaux sont trop petits pour être étudiés et toutes les données obtenues concernent uniquement les cônes parafovéaux. Chaque cône, au moins extrafovéal, ne contient apparemment qu'un seul des pigments ou majoritairement l'un d'entre eux.
Méthodes modernes pour étudier les photopigments des cônes
Densitométrie des cellules rétiniennes
→ densitométrie, peut être la suivante :
Si les premières tentatives pour trouver trois pigments et, comme prévu, trois types de cônes ont été réalisées par Rushton, qui a perfectionné la technique Densitométrie pour mesurer les coefficients d'absorption de la lumière avec différentes longueurs d'onde - dans la couche de photorécepteurs de la rétine, et il a été démontré que les anomalies de couleur manquent de l'un des pigments trouvés chez les personnes ayant une vision normale : « erythrolab » (maximum environ 585 nm.) dans protanope et "chlorolab" ( maximum environ 540 nm.) - dans le deutéranope, désormais à l'aide de méthodes de dynestométrie spéciales utilisant des densitomètres modernes, les scientifiques sont capables de déterminer le fonctionnement des cônes et des bâtonnets dans un état d'activité normale et de diagnostiquer leurs maladies.
voir également
Sources
Littérature
- Khokhlova T.V. Idées modernes sur la vision des mammifères // Journal of General Biology. Volume 73, 2012. N° 6, novembre-décembre. Page 418-434.