vizuális pigmentek. fotorecepció. A szemgolyó héjai közül melyik átlátszó elöl. Az oldalsó geniculate test egy szerkezet
VIZUÁLIS PIGMENTEK(lat. pigmentum festék) - a retina fotoreceptorainak fényérzékeny pigmentjei. 3. A fényimpulzus energiáját érzékelve összetett fotokémiai cikluson mennek keresztül. transzformációk, amelyek eredményeként a retina különálló, 3. p.-ot tartalmazó vizuális receptora (kúp vagy rúd) gerjesztett állapotba kerül és a kapott információt a látóideg mentén továbbítja a c. n. Val vel. A retina vizuális sejtjei fotoreceptor membránjának fő szerkezeti és funkcionális részeként 3. o. kulcsszerep a látás mechanizmusaiban (lásd).
Nómenklatúra és szerkezet vizuális pigmentek. A gerincesek és gerinctelenek összes vizsgált 3. oldala a vízben oldhatatlan membránfehérje opszin és a hozzá kapcsolódó kromofor (retina) komplexei. A retinális vagy az A-vitamin-aldehid két formában létezhet, a retina1 és a retina2 formájában.
A kromofor 3. természeténél fogva az elem két osztályra oszlik - a retinális1-et tartalmazó rodopszinokra (lásd.) és a retinális2-t tartalmazó porfiropzinokra. A rodopszinok minden szárazföldi és tengeri állat szemének retinájában, a porfiropzinok az édesvízi állatok szemének retinájában találhatók. Egyes halak és kétéltűek egyidejűleg retinális 3 p.-t tartalmaznak! és retina. Vannak kísérletek a 3. p. osztályozására a retina rúdjaira vagy kúpjaira jellemző opszinok különbségei alapján. Például a rodopszin a retina1 és a rúd-opszin komplexe, a jodopszin a retina1 és a kúpos opszin komplexe, a porfiropzin a retina2 és a rúd-opszin komplexe, a retina-kúp opszin komplex cianopzint képez. A 3. tételeket azonban rendkívül nehéz opszinok alapján osztályozni, mivel legalább öt különböző opszin létezik.
Az összes ismert 3. tétel közül a bika, béka és tintahal szeméből izolált rodopszinokat vizsgálták a legteljesebben. A könyörgésük. 30-40 ezres nagyságrendű tömeg (tömeg), minden molekula kb. 400 aminosav és egy kromofor. Ezenkívül az oligoszacharid lánc a 3. p. molekulájában szerepel: 3 glükózamin gyök, 2 mannóz, 1 galaktóz. A lipidek (ch. arr. foszfolipidek) erős komplexet alkotnak a 3. molekulával. Az alapvető spektrális tulajdonságaik megőrzése mellett (lásd Spektrális analízis) a lipidek nélküli tételek 3. elveszítenek számos funkcionálisan fontosat, például a helyreállítási képességüket.
A tiszta retina sárga színű, abszorpciós spektruma maximuma 370 nm tartományban van. Az Opsin színtelen, az abszorpciós maximum az ultraibolya tartományban van (kb. 280 nm). A rodopszin molekula színe vöröses-rózsaszín, maximális abszorpciós spektruma kb. 500 nm. A komplex képződése során bekövetkező ilyen erős spektrális eltolódás oka (370-ről 500 nm-re - az úgynevezett batokróm eltolódás) még nem kapott egyértelmű magyarázatot.
A rodopsinek és porfiropzinok abszorpciós spektrumának maximumai a látható spektrum meglehetősen széles tartományát ragadják meg - rodopsinek esetében 433-562 nm, porfiropzinok esetében pedig 510-543 nm. Ha a porfiropzinok közé tartoznak a béka, ponty és édesvízi teknős ebihalának 3 p.-kúpjai is, azaz cianopszin maximális abszorpciós spektrummal 620 nm-en, akkor ez a tartomány még szélesebb. A mikrospektrofotometriás módszerek fejlődése lehetővé tette számos típusú egyetlen fotoreceptor sejt abszorpciós spektrumának meghatározását állatokban és emberekben. A kapott adatok szerint az emberi retina 3 p.-jának a következő abszorpciós spektrum maximumai vannak: pálcikák 498, kékre, zöldre és vörösre érzékeny kúpok - 440, 535 és 575 nm.
A 3. o. tanulmányozása németül indult. H. Muller kutató, aki 1851-ben leírta, hogy a béka szeméből kivont rózsaszínes-lila retina a fényben először sárgássá, majd fehéressé válik. 1877-ben F. Boll is leírta ezt a jelenséget, és arra a következtetésre jutott, hogy a retina látósejtjeiben valamilyen vörös fényre érzékeny anyag található, és ennek az anyagnak az elszíneződése a látás mechanizmusával függ össze. A 3. tétel tanulmányozásában nagy érdem Kuhne (W. Kuhne, 1877), Kromnak sikerült azonosítania és részletesen tanulmányoznia a 3. tételt. Az általa kivont 3. p.-ot vizuális lilának nevezte, megállapította fehérjetermészetét, megvizsgálta egyes spektrális tulajdonságait és fototranszformációit, felfedezte a 3. p.-nek a sötétben való regenerálódási képességét. Amer. nagyban hozzájárult a 3. p. biokémikus és fiziológus J. Wald.
Vizuális pigmentek fototranszformációi. Fény hatására a 3. p.-on fotokémiai körfolyamat megy végbe bennük. transzformációk, amelyek a retina cisz-transz izomerizációjának elsődleges fotokémiai reakcióján alapulnak (lásd Izomerizmus). Ebben az esetben a kromofor és a fehérje közötti kötés megszakad. A 3. p. átalakulásainak sorrendje a következőképpen ábrázolható: rodopszin (a kromofor cisz formában van) -> prelumirhodopsin -> lumirhodopsin -> metarhodopsin I -> metarhodopsin II -> opsin protein -> kromofor transz formában. Az enzim - retinol-dehidrogenáz - hatására ez utóbbi A-vitaminba megy át, amely a rudak és kúpok külső szegmenseiből a retina pigmentrétegének sejtjeibe kerül. Amikor a szem elsötétül, a regeneráció 3. p. következik be, a vágás végrehajtásához az A-vitamin cisz-izomerjének jelenléte, amely a kromofor képződésének kiindulási terméke (A-vitamin-aldehid) szükséges. Az A-vitamin hiányával vagy hiányával a szervezetben a rodopszin képződése megszakadhat és kialakulhat a szürkületi látás zavara, az ún. éjszakai vakság (lásd Hemeralopia). A rodopszin fototranszformációjának folyamatában a lumirhodopszin metarodopszin I-re való átmenetének szakaszában a receptorsejtben fényes villanás hatására következik be az ún. korai (rövid látenciájú) receptorpotenciál. Ugyanakkor nem vizuális jel, bár az egyik tesztként szolgálhat a 3. p.-nek a fotoreceptor membránjában bekövetkező átalakulási mechanizmusának tanulmányozására. Funkcionális érték az ún. késői receptorpotenciál, a látens periódus to-rogo (5-10 msec) arányos a metarodopszin II képződésének idejével. Feltételezhető, hogy a metarodopszin I metarodopszin II átalakulásának reakciója biztosítja a látszatot. vizuális jel.
Mivel a 3. tárgyak folyamatosan elszíneződnek a fényben, léteznie kell a folyamatos helyreállításuknak. Egyesek rendkívül gyorsak (fotoregeneráció), mások meglehetősen gyorsak (biokémiai, regenerációs, Sötét), mások lassúak (3. p. szintézise A látósejtben a fotoreceptor membrán állandó megújulása során). A fotoregenerációnak van fiziol értéke, gerinctelen állatoknál (például at lábasfejűek- tintahal, polip). A biokémiai mechanizmusban. gerinceseknél fontos szerepe van az izomeráz enzimnek (lásd), amely biztosítja a transz-retinális (vagy transz-A-vitamin) izomerizációját ismét cisz izomer formává. Nincs azonban végleges bizonyíték egy ilyen enzim létezésére. Ugyanez a reakció, amikor egy 3. p molekula keletkezik a retina 11-cisz-izomerje és az opszin jelenlétében a rendszerben, könnyen, energiafelhasználás nélkül megy végbe. Megállapítottuk, hogy a fehérített rodopszin képes a foszforilációs reakcióra (lásd); Feltételezzük, hogy ez a reakció a látósejt fényadaptációjának mechanizmusának egyik láncszeme.
Bibliográfia: Ackerman Yu. Biofizika, ford. angolból, M., 1964; Willie K. és Det e V. Biology, ford. angolból, M., 1974, bibliográfia; Konev S. V. és Volotovsky I. D. Bevezetés a molekuláris fotobiológiába, p. 61, Minszk, 1971; Osztrovszkij M. A. és Fedorovich I. B. Fotoindukált változások a fotoreceptor membránjában, a könyvben: Structure and functions of biol, membránok, szerk. A.S. Troshina és mások, p. 224, M., 1975, bibliogr.; Érzékszervi rendszerek élettana, szerk. G. V. Gershuni, 1. rész, p. 88, L., 1971; Vizuális pigmentek biokémiája és élettana, szerk. szerző: H. Langer, B. a. o., 1973; Érzékélettani kézikönyv, szerk. írta: H. A. R. Jung a. o., v. 7, 1-2. pont, B., 1972.
M. A. Osztrovszkij.
vizuális pigment A retina fotoreceptorainak fényérzékeny membránjának szerkezeti és funkcionális egysége (lásd: Fotoreceptorok) - rudak és kúpok. Z. o.-ban a vizuális észlelés első szakaszát hajtják végre - a látható fény kvantumainak elnyelését. A Z. molekula (móltömeg körülbelül 40 000) egy fényelnyelő kromoforból és opszinból, fehérje és foszfolipidek komplexéből áll. Az összes Z. p. kromoforja az A 1 vagy A 2 vitamin aldehidje - retina vagy 3-dehidroretinális. Kétféle opszin (rúd és kúp) és kétféle retina párban kombinálva 4 típusú z.p. nm), jodopszin (562 nm), porfiropzin (522 nm) és cianopszin (620 nm). A látás mechanizmusának elsődleges fotokémiai láncszeme (Lásd Vision) a retina fotoizomerizációjában áll, amely fény hatására görbült alakját laposra változtatja. Ezt a reakciót sötét folyamatok láncolata követi, amely vizuális receptor jel megjelenéséhez vezet, amely szinaptikusan továbbítódik a retina következő idegelemeihez - bipoláris és horizontális sejtekhez. Megvilágított.:Érzékszervi rendszerek élettana, 1. rész, L., 1971, p. 88-125 (élettani kézikönyv); Wald G., A vizuális gerjesztés molekuláris alapja, "Természet", 1968, v. 219. M. A. Osztrovszkij.
Nagy Szovjet Enciklopédia. - M.: Szovjet Enciklopédia. 1969-1978 .
Nézze meg, mi a "Visual pigment" más szótárakban:
Szerkezetileg működőképes. fényérzékeny egység. rúd és kúp fotoreceptor membránok a retinában. A 3. p. molekula egy fényelnyelő kromoforból és egy fehérje és foszfolipidek komplexének opszinjából áll. A kromofort az A1-vitamin aldehid képviseli ... Biológiai enciklopédikus szótár
A rodopszin (vizuális lila) a fő vizuális pigment az emberi és állati retina pálcáiban. Komplex fehérjék kromoproteinekre utal. A különböző biológiai fajokra jellemző fehérjemódosítások jelentősen eltérhetnek ... Wikipédia
VIZUÁLIS(E) PIGMENT(EK)- Lásd a fotopigmentet... Szótár a pszichológiában
A rudak belsejében található retina pigment, amely magában foglalja a retina (retina) A-vitamint és fehérjét. A rodopszin jelenléte a retinában szükséges a normál látáshoz gyenge fényben. A fény hatására ...... orvosi kifejezések
RHODOPSIN (RODOPSIN), LILA VIZUÁL- (vizuális lila) retina pigment a pálcikák belsejében, amely retina (retinális) A-vitamint és fehérjét tartalmaz. A rodopszin jelenléte a retinában szükséges a normál látáshoz gyenge fényben. alatt…… Magyarázó orvosi szótár
- (vizuális lila), fényérzékeny. komplex fehérje, pálcikasejtek vizuális pigmentje gerincesek és emberek retinájában. Egy fénykvantumot elnyelve (abszorpciós maximum kb. 500 nm), az R. szétesik és gerjesztést okoz ... ... Természettudomány. enciklopédikus szótár
- (vizuális pigment), gerincesek retinájának és gerinctelenek látósejtjeinek fényérzékeny rúdfehérje. R. glikoprotein (mol.m. kb. 40 ezer; polipeptid lánc 348 aminosavból áll), amely ... ... Kémiai Enciklopédia
- (a görög rhódon rose és ópsis látás szóból) vizuális lila, gerincesek (egyes halak és kétéltűek kivételével a fejlődés korai szakaszában) és gerinctelenek retinarudainak fő vizuális pigmentje. A vegyszer szerint ...... Nagy Szovjet Enciklopédia
- (vizuális bíbor), fényérzékeny komplex fehérje, gerinceseknél és embereknél a retina rúdsejtjeinek fő vizuális pigmentje. Egy fénykvantumot elnyelve (a maximális abszorpció körülbelül 500 nm) a rodopszin lebomlik és ... ... enciklopédikus szótár
Fő cikk: Rudak (retina) A rodopszin (egy elavult, de még mindig használt név: vizuális lila) a fő vizuális pigment. Tengeri gerinctelenek, halak, szinte minden szárazföldi élőlény szem retinájának rúdjaiban található ... ... Wikipédia
25-10-2014, 13:04
A fénykvantumokat speciális molekulák - vizuális fotopigmentek - abszorbeálják a receptorokban. A vizuális pigmenteket egymástól függetlenül fedezték fel F. Boll és W. Kuehne német fiziológusok 1877-1879-ben. F. Boll észrevette, hogy a béka vizuális üvegéből izolált retina először vörösnek tűnik, majd a fényben elhalványulva sárgává, végül teljesen színtelenné válik.
W. Kuehne azt találta, hogy egy állat normálisan működő szemében a retina színe intenzív fénnyel történő megvilágítás után helyreáll, ha az állatot ismét sötétbe helyezik. Ezen megfigyelések alapján arra a következtetésre jutottak, hogy a szem retinájában egy fényérzékeny anyag (pigment) található, melynek koncentrációja fényben csökken, sötétben pedig helyreáll.
vizuális pigmentek A kromolipoproteinek összetett molekulái, amelyek gerincesekben és gerinctelenekben két fő részből állnak: egy kromoforból (a molekula színező része, amely megvilágítva határozza meg a receptor színét) és egy fehérjéből (opszin). A kromofor meghatározza a pigmentben a fényelnyelés maximumát és intenzitását, és alkoholok - A1 és A2 vitaminok - aldehidje.
Ezeknek az aldehideknek a neve retina-1 és retinal-2. Egy további kettős kötés jelenléte a retina-2-ben az abszorpciós spektrum maximumának a hosszú hullámhosszú tartományba való eltolódásához vezet. A retina az opszinhoz kötődik, egy fehérjemutációhoz, amely csak a fotoreceptorokban található. Az opszin rúd, a scotopsin és a kúpok, a fotopszin, izolálva vannak. A Scotopsin 230-270 aminosavból áll, amelyek szekvenciáját még nem határozták meg.
A fotopszinról keveset tudunk: csak a spirális konfigurációjáról van adat. A pigmentmolekula tényleges fényelnyelő része, a retina egyfajta karotinoid pigment, a legelterjedtebb a fotoszintetikus folyamatokban. növények és állatok szövetei.
A pigmentek spektrális jellemzőit az egyik retina és az opszinok egyik vagy másik változatának kombinációja határozza meg, amelyek sokfélesége meghatározza a vizuális pigmentek sokféleségét a különböző állatokban. Így a scotopsin és a retina-1 kombinációja egy rúdpigment - rodopszin képződéséhez vezet, amelynek abszorpciós maximuma a = 500 nm-nél van, és a retina-2-vel való kombinációja egy másik pálcika pigmentet képez - porfiropzin, amelynek abszorpciós maximuma at? =525 nm. A retina-1 kombinációi a másfajta az emberek és majmok retinájában lévő fotopszinek kúppigmenteket alkotnak, amelyek abszorpciós maximuma a? = 445 nm, ? = 535 nm és? \u003d 570 nm, ezeket cianolabnak, klorolabnak és eritrolabnak nevezik.
A pigmentek fényelnyelő tulajdonságait pontosan az iopszin kromofor kölcsönhatása határozza meg: a kromofor és az opszin abszorpciós maximumai külön-külön is a régióban lokalizálódnak? \u003d 278 nm (retina-1) és? = 378 nm (scotopsin), míg vegyületük - rodopszin - abszorpciós maximuma a? = 500 nm. Az abszorpciós maximum a pigment spektrális jellemzőinek egyik paramétere, amely általában meghatározza a fénykvantumok (fotonok) pigment általi abszorpciójának hatékonyságát. különböző hosszúságú hullámok (3.1.4a ábra).
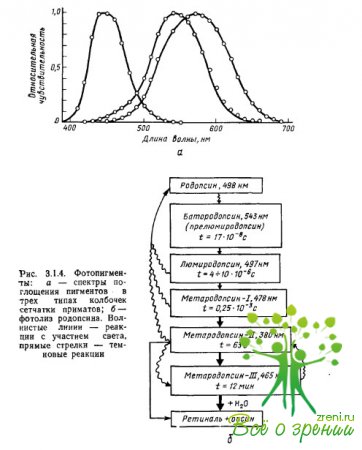
A pigmentek abszorpciós spektrumának első mérését majmok és emberek retinájának egyes kúpjaiban Brown, Wald és Marx és munkatársai végezték, speciálisan erre a célra kifejlesztett mikrospektrofotometriás módszerrel. A retina preparátumot mikroszkóp alá helyezték, ami vizuális ellenőrzés mellett lehetővé tette, hogy egy vagy olyan hullámhosszú fénysugarat a pigmentet tartalmazó fotoreceptor külső szegmensére fókuszáljon.
Ezután minden hullámhosszon külön-külön megmértük a kúpon áthaladó és általa el nem nyelt fény mennyiségét (a kúpba juttatott és az általa átbocsátott fénymennyiség különbsége jellemezte az elnyelés hatékonyságát adott hullámhosszúságú fotonok a pigment által). A méréseket megismételtük az intenzív megvilágítás hatására bekövetkező pigment fakulást követően (referencia mérések). A második méréssorozatnak az elsőből való kivonása eredményeként a pigment differenciális abszorpciós spektrumát kaptuk a sugárzási hullámhosszon mért fényérzékenység függvényében.
ábrából. A 3.1.4a ábrán látható, hogy a pigmentek széles sávú, erősen átfedő abszorpciós spektrummal rendelkeznek. Minden fotopigment rendelkezik a karotinoidokra jellemző további abszorpciós maximummal (az úgynevezett nis-csúcs" vagy "?-csúcs"), amely a fő maximum 2/3-ával egyenlő. Azoknál a pigmenteknél, amelyek fő abszorpciós maximumai a spektrum rövid és közepes hullámhosszú részében találhatók, a cisz-csúcs az ultraibolya tartományba esik, és nem befolyásolja a látható spektrum sugárzásának elemzését. Ha továbbá a pigmentek spektrális érzékenységének függvényeinek megszerkesztésekor a hullámhossz-skálát (abszcissza tengely) frekvenciaskálává alakítjuk, akkor az összes pigment abszorpciós spektruma, beleértve a különböző opszinokat is, de ugyanaz a retina (például retina). -1), ugyanolyan és azonos alakú lesz.
Az adatoknak ezzel a reprezentációjával, amelyet először Dartnall (1953) javasolt a retina-1 alapú pigmentekre, a különböző fotopigmentek abszorpciós spektruma csak a maximum pozíciójában tér el. És akkor, ismerve a pigment abszorpciós maximumát, vissza tudja állítani abszorpciós spektrumának alakját, először a „sugárzási frekvencia – érzékenység”, majd a c = ?f arány és a „hullámhossz – érzékenység” koordinátákban.
Ezt a módszert, amellyel egy ismeretlen pigment abszorpciós spektrumát csak a maximum adataiból állítják helyre, nomogram módszernek nevezik. A nomogram módszert széles körben alkalmazzák a színlátás neurofiziológiájában is az egyik vagy másik színérzékeny neuronhoz konvergáló fotodetektorok számának és típusának problémájának megoldása során: egy neuron spektrális érzékenységi függvényét összehasonlítják egyik vagy másik nomogrammal (ill. kombinációjuk).
A fotopigmentek sokfélesége az állatvilágban korrelál a környezeti élőhelyek sokféleségével. Halaknál és kétéltűeknél a retina egyaránt tartalmazhat rodopszint és porfiropzint, amelyek aránya az egyed élete során változik; az anadrom halakban a rodopszin porfiropzinnal való helyettesítése megelőzi a sós vízből édesvízbe való átmenetet és fordítva. A pigmentkészlet kapcsolata az élőhelyökológiával nem merev: a pigment összetétel változását szezonális változások és cirkadián ritmusok okozhatják.
Fotoizomerizáció.
A fotopigmentek fényre adott reakcióját az elnyelt kvantumok száma határozza meg, függetlenül a hullámhossztól és a sugárzási energiától: például a rodopszin által elnyelt kvantumok. 10 fénymennyiség? \u003d 430 nm ugyanazokat a szerkezeti változásokat okozza benne (3.1.46. ábra), valamint 10 elnyelt fénykvantum? = 500 nm.
A pigment spektrális szelektivitását (abszorpciós spektrumát) pedig csak a látható spektrum különböző részeiből származó fénykvantumok elnyelésének eltérő valószínűsége határozza meg. Így a vizuális pigmentek és végső soron a fotoreceptorok "fényfotonok számlálóiként" működnek, és csak a különböző hullámhosszú fotonok elnyelésének valószínűségében különböznek egymástól.
Amikor egy rodopszinmolekula 1 kvantum fényt nyel el, a retina egyik molekulájának térbeli konfigurációja megváltozik - fotoizomerizáció, amely abból áll, hogy a retina 11 cm-es formája teljesen átalakul (3.1.4b. ábra). .
A fotoizomerizációt fotokémiai reakciók köre kíséri, amelynek dekódolásáért G. Wald 1930-ban Nobel-díjat kapott. A fotoizomerizáció végtermékei teljesen traxretinál és opszin. A metarodopszin-II lépésig minden közbenső reakció reverzibilis: e reakciók termékei reverz gran-retinális izomerizációval visszaalakíthatók rodopszinná.
Ez azt jelenti, hogy a pigmentmolekulában nem történik jelentős konformációs változás a metarodopszin-II előtt. A metarodopszin-II stádiumában a retina leválik a molekula fehérjerészéről. Az izomerizáció összes szakasza közül csak az elsőnek - a rodopszin prelumirhodopszinná (bathorhodopsin) való átalakulásának - van szüksége fényre. Az összes többi reakció sötétben megy végbe. A fotoizomerizáció hasonlóan megy végbe a kúpos pigmentekben.
Így a fotoizomerizáció a fotorecepció molekuláris kiváltó (indító) mechanizmusa, amely a vizuális pigmentmolekula térbeli konfigurációjának megváltoztatásából áll. Ez utóbbi egyértelművé teszi, hogy a természet miért választotta a karotinoidokat vizuális pigmentként: a kettős kötések jelenléte a retinában meghatározza annak izomerizációjának lehetőségét és; magas reaktivitása. Minden kettős kötés körül a molekula egyes részei elfordulhatnak és elhajolhatnak a térben. Ez a forma a legkevésbé stabil és a legérzékenyebb a hőmérsékletre és a fényre. A természetben megtalálható különböző fajták retina - 7, 9, 11, 13-cisz-forma, 9,13-dicis-forma, de vizuális pigmentekben a retina mindig 11-cisz-formában van.
A fotoreceptorokban a pigment reszintézis folyamatosan megy végbe világosban és sötétben a retinén izomeráz enzim részvételével, és a retinol fordított izomerizációjából áll a transzformációból a 11-cisz formába. A kúpokban ez az enzim magukban a receptorokban, pálcikákban, a pigment epitéliumban található, ahová az opszinról való leválás után a transz-retinol áthalad, és ahol izomerizálódik 11-cisz formává. Ebben az esetben az A-vitamin retináját újra felhasználják, és a fehérjét a belső szegmens riboszómáiban szintetizálják.
Feltételezzük, hogy a 11-cisz-retinál és az opszin rekombinációja spontán módon megy végbe. A pigment epitélium körülbelül 11%-ban A-vitamint (retinolt) tartalmaz a teljes szem 13%-ából. Az A-vitamin a májban úgy képződik, hogy a táplálékban lévő karotinoid láncát két részre bontják és vizet adnak hozzá.
Az A-vitamint egy speciális retinol-kötő fehérje szállítja a pigmenthámba, ahol a retinává oxidálódik. Az A-vitamin hiánya esetén az úgynevezett "éjszakai vakság" következik be - az abszolút fényérzékenység csökkenése, ami különösen az alkonyatkor érinti a látást, amely egy szabad állapotban instabil fehérje megsemmisülése miatt tartós vaksággá válik. Ezért az "éjszakai vakság" betegsége esetén ajánlatos A-provitamint - béta-karotint tartalmazó sárgarépát fogyasztani.
Minden fotoreceptor csak egy fotopigmentet tartalmaz, amelyet egy vagy másik abszorpciós spektrum jellemez. Ebben a tekintetben rövid, közepes és hosszú hullámhosszú kúpokat izolálnak, amelyek pigmenteket tartalmaznak a főemlősökben, abszorpciós maximummal 445 nm-en, 535 nm-en és 570 nm-en. Nál nél édesvízi hal a hosszú hullámhosszú kúp 620 nm-en abszorpciós maximummal rendelkező pigmentet tartalmaz, ami korrelál a fény spektrális eloszlásának alakjával az élőhelyükön.
Ezeket a kúpokat néha kékre, zöldre és vörösre érzékeny receptoroknak nevezik, a háromkomponensű Young-Helmholtz hipotézisig visszanyúló hagyományt követve. De a 20. század második fele óta, amikor kiderült, hogy önmagában egyetlen fotoreceptor sem végez spektrális elemzést, hanem csak az elnyelt fotonok számára reagál, függetlenül a sugárzás hullámhosszától, a receptorok ezen színneveit csak metaforikusan használják. érzék.
A rudak besorolása például a békáknál „zöldre” és „pirosra” bizonyos esetekben nem a bennük lévő pigment spektrális érzékenységén alapul, hanem a receptortestek színén az áteresztett fényben. A "zöld" rudak tehát 432 nm-en abszorpciós maximummal rendelkező pigmentet tartalmaznak, ezért fehér fénnyel megvilágítva elnyelik a rövidhullámú sugárzást, minden más hullámhosszon áthaladva, vizuálisan szabályozva pedig zöldesnek tűnnek.
A halak retinájában összefüggést figyeltek meg a kúp hossza és a benne lévő pigment spektrális típusa között: közülük a legrövidebb rövidhullámú, a leghosszabb pedig hosszú hullámú pigmentet tartalmaz. Eberle felvetette, hogy ez lehet a kromatikus aberráció elleni küzdelem egyik mechanizmusa. A kromatikus aberráció miatt a középhullámú sugárzás éles képe mélyebben (az objektívtől távolabb) helyezkedik el, mint a rövidhullámú sugárzás éles képe, a hosszúhullámú sugárzás fókusza pedig még messzebbre lokalizálódik.
Elegendő a rövid, közepes és hosszú hullámhosszú receptorokat megfelelő szinten elrendezni, hogy a spektrális sugárzás minden része egyformán élesen fókuszáljon a receptorokra. Talán ez magyarázza azt a tényt, hogy a halak retinájában a rövid szegmensű egykúpok rövid hullámhosszú pigmentet tartalmaznak, míg a kettős kúpban a leghosszabb szegmensben a hosszú, a rövidebb pedig a rövid hullámhosszú pigmentet. vagy közepes hullámhosszú pigment. Az egyenlő hosszúságú szegmensekkel rendelkező kettős kúpok ugyanazt a pigmentet tartalmazzák.
ωπς - szem) - gyakori név emberek és egyes emlősök számos vizuális pigmentje. Ezek a pigmentek a retina kromoforjához kapcsolódó fehérjemolekulából állnak. A retina kúpjaiban található, és színlátást biztosít, ellentétben egy másik vizuális pigmenttel - a rodopszinnal, amely felelős az alkonyi látásért.Megállapítást nyert, hogy a vizuális pigmentek összetétele opszinokat tartalmaz. A különböző opszinok aminosav-molekuláris összetételükben különböznek, és kissé eltérő hullámhosszúságú fényt nyelnek el, mint például a retinához kötött molekulák.
A kúpos pigment létezését először George Wald fedezte fel (közvetve), ő adta a nevét. jodopszin. . Ezekért a munkáiért 1967-ben fiziológiai vagy orvosi Nobel-díjat kapott.
Még Helmholtz is úgy gondolta, hogy a "látóideg-szálak terminális apparátusában" (a retina fotoreceptoraiban) háromféle fotokémiailag lebomlott anyagnak kell lennie, amelyek eltérő érzékenységgel rendelkeznek Különböző részek spektrum. .
A modern tudományban uralkodó háromkomponensű látáselmélet szerint ennek a pigmentnek három típusa van, a retina pedig háromféle kúpot tartalmaz, amelyek érzékenyek a kék, zöld és vörös fényre. Ennek megfelelően a jodopszin, amely a szem kúpjaiban található, három pigmentből áll - klorolab, eritrolab és cianolab; az első a sárga-zöld (450-630 nm abszorpciós sáv), a második - sárga-piros (500-700 nm) és a harmadik - kék-zöld (500-700 nm) sugarakat nyeli el. spektrum.
A vizuális pigmentek típusai
Az első kísérletet három pigment és, ahogy az várható volt, háromféle kúp megtalálására (a háromkomponensű látás hipotézisének feltételezései alapján, hogy minden kúp csak egy pigmentet tartalmaz) Rushton végezte, aki tökéletesítette az in vivo denzitometriás technikát. különböző hullámhosszú fény abszorpciós együtthatóinak mérése - a rétegben a retina fotoreceptoraiban. Kimutatták, hogy a színeltérésekből hiányzik a normál látású emberekben jelenlévő pigmentek egyike: az "erythrolab" (maximum kb. 585 nm) a protanopban és a "chlorolab" (maximum kb. 540 nm) a deuteranopban.
Meg kell jegyezni, hogy a „vevő” kifejezés használatakor bizonyos esetekben az azonos spektrális érzékenységű fotoreceptorok teljes készletét egynek tekintjük. vevő; más esetekben az a kérdés, hogy mindegyik foveális kúp tartalmaz-e hármat vevő vagy csak egy. Ugyanakkor nem sérül a "vevő" fogalmának szigorúsága, amely ebben az esetben nem tartalmaz konkrét morfológiai jellemzőket.
A következő lépés ebben az irányban az egyes ember- és makákókúpokban található fotopigmentek vizsgálata volt. A foveális kúpok mérete túl kicsi ahhoz, hogy vizsgálat tárgyát képezze, és minden kapott adat csak a parafoveális kúpokra vonatkozik. Minden kúp, legalábbis az extrafovealis, úgy tűnik, csak egy pigmentet tartalmaz, vagy túlnyomórészt egyet.
Modern módszerek a kúpos fotopigmentek tanulmányozására
Retina sejt denzitometria
→ denzitometria, a következő lehet:
Ha az első kísérletet három pigment és a várakozásoknak megfelelően háromféle kúp megtalálására Rushton végezte, aki tökéletesítette a technikát Denzitometria a különböző hullámhosszú fény abszorpciós együtthatóinak mérésére - a retina fotoreceptorok rétegében, és kimutatták, hogy a színrendellenességekből hiányzik a normál látású emberek egyik pigmentje: "erythrolab" (maximum kb. 585 nm) a protanopban és " chlorolab" (maximum körülbelül 540 nm.) - a deuteranopban, most a modern denzitométereket használó speciális disztometriai módszerek segítségével a tudósoknak sikerül meghatározniuk a kúpok és rudak munkáját normál aktivitású állapotban, és diagnosztizálni betegségeiket.
Lásd még
Források
Irodalom
- Khokhlova T.V. Modern ötletek az emlősök látásáról // Journal of General Biology. 73. évfolyam, 2012. 6. szám, november-december. oldal 418-434.