görsel pigmentler Derinin epidermisinin ilkel bölgesi katmanlardan oluşur. Gündüz ve gece görüşü
Görsel pigmentler, dış bölümlerin zarlarında yoğunlaşmıştır. Her çubuk yaklaşık 108 pigment molekülü içerir. Dış zara bağlı olmayan birkaç yüz ayrı diskte (maymun değneklerinde yaklaşık 750) organize edilirler. Konilerde pigment, fotoreseptörün dış hücre zarının devamı olan özel pigment kıvrımlarında bulunur. Pigment molekülleri, tüm disk proteinlerinin yaklaşık %80'ini oluşturur. Görsel pigmentler, dış segmentin zarlarında o kadar yoğun bir şekilde paketlenir ki, çubuktaki iki görsel pigment molekülü arasındaki mesafe 10 nm'yi geçmez. Bu tür yoğun paketleme, bir fotoreseptör hücre katmanından geçen bir ışık fotonunun yakalanma olasılığını artırır. Şu soru ortaya çıkıyor: Işık görsel pigmentler tarafından emildiğinde sinyaller nasıl ortaya çıkıyor?
Görsel pigmentler tarafından ışık emilimiIşık çubuk pigmenti, rodopsin tarafından emildiğinde meydana gelen olaylar, psikofizyolojik, biyokimyasal ve moleküler teknikler kullanılarak incelenmiştir. Görsel pigment molekülü iki bileşenden oluşur: opsin adı verilen bir protein ve retina adı verilen bir kromofor, 11-cis-vitamin A-aldehit (Şekil 1). Kromoforun bileşiğe renk veren kimyasal bir grup içerdiği açıklığa kavuşturulmalıdır. Pigmentlerin absorpsiyon kapasitesinin nicel özellikleri, spektrofotometri kullanılarak incelenmiştir. Çubukların görsel pigmenti olan rodopsin, farklı dalga boylarında ışıkla aydınlatıldığında, yaklaşık 500 nm dalga boyuna sahip mavi-yeşil ışık en iyi şekilde emildi. Benzer bir sonuç, mikroskop altında tek bir çubuğun farklı dalga boylarında ışık huzmeleri ile aydınlatılmasıyla da elde edildi. Ortaya çıktı ilginç bağımlılık Rodopsinin absorpsiyon spektrumu ile alacakaranlık ışığını algılamamız arasında. İnsanlar üzerinde yapılan nicel psikofiziksel çalışmalar, yaklaşık 500 nm dalga boyuna sahip mavimsi yeşil gün ışığının karanlıkta alacakaranlık ışığının algılanması için optimal olduğunu göstermiştir. Çubukların aktif olmadığı ve sadece konilerin kullanıldığı gün boyunca, konilerin absorpsiyon spektrumuna karşılık gelen kırmızı renge en duyarlıyız (bundan daha sonra bahsedeceğiz).
Bir foton rodopsin tarafından absorbe edildiğinde, retinal fotoizomerizasyona uğrar ve 11-cis'ten trans konfigürasyonuna geçer. Bu geçiş çok hızlı gerçekleşir: yaklaşık 10-12 saniye içinde. Bundan sonra, pigmentin protein kısmı da bir dizi ara ürünün oluşumuyla birlikte bir dizi dönüşümsel değişikliğe uğrar. Protein parçasının konformasyonlarından biri olan metarhodopsin II, sinyal iletimi için en önemlisidir (bunu bu bölümde daha sonra tartışacağız). Şekil 2, aktif rodopsinin renginin giderilmesi ve rejenerasyonu sırasındaki olayların sırasını göstermektedir. 1 ms sonra Metarhodopsin II oluşur. Parçalanmasından sonra pigment rejenerasyonu birkaç dakika içinde yavaş yavaş gerçekleşir; bu, retinanın fotoreseptörlerden pigment epiteline taşınmasını gerektirir.
Rodopsin'in yapısıMoleküler düzeyde, opsin proteini, her biri 20-25 amino asitten oluşan 7 hidrofobik bölge oluşturan ve 7 transmembran sarmal oluşturan 348 amino asit kalıntısından oluşur. Molekülün N-ucu hücre dışı boşlukta (yani çubuk diskin içinde) bulunur ve C-ucu sitoplazmada bulunur.
Karanlıkta, 11-cis-retinal, opsin proteinine sıkıca bağlıdır. Foton yakalama, retina tahtındaki tüm cis retinalin izomerizasyonuna yol açar. Bu durumda, opsin all-tron-retinal kompleksi hızla, opsin ve all-tron retinal'e ayrışan metarhodopsin II'ye dönüşür. Rodopsin rejenerasyonu, fotoreseptörler ve pigment hücrelerinin etkileşimine bağlıdır. Metarhodopsin II, ikinci haberci sistemini aktive eder ve sürdürür.
Retina, yedinci transmembran segmentinde bulunan bir lizin tortusu yoluyla opsine bağlanır. Opsin, adrenerjik ve muskarinik reseptörler gibi metabotropik aracı reseptörleri de içeren 7 transmembran alanına sahip bir protein ailesine aittir. Rodopsin gibi, bu reseptörler G-protein aktivasyonu yoluyla ikinci habercilere sinyal verir. Rodopsin karanlıkta oldukça kararlıdır. Bayor, rodopsin molekülünün kendiliğinden termal izomerizasyonunun yaklaşık 3000 yıl veya fotoizomerizasyondan 1023 daha fazla sürdüğünü hesapladı.
görsel pigmentler
görsel pigmentler
Rodopsin'in yapısı
Koniler ve renkli görme
renk körlüğü
Edebiyat
görsel pigmentler
Görsel pigmentler, dış bölümlerin zarlarında yoğunlaşmıştır. Her çubuk yaklaşık 108 pigment molekülü içerir. Dış zara bağlı olmayan birkaç yüz ayrı diskte (maymun değneklerinde yaklaşık 750) organize edilirler. Konilerde pigment, fotoreseptörün dış hücre zarının devamı olan özel pigment kıvrımlarında bulunur. Pigment molekülleri, tüm disk proteinlerinin yaklaşık %80'ini oluşturur. Görsel pigmentler, dış segmentin zarlarında o kadar yoğun bir şekilde paketlenir ki, çubuktaki iki görsel pigment molekülü arasındaki mesafe 10 nm'yi geçmez. Bu tür yoğun paketleme, bir fotoreseptör hücre katmanından geçen bir ışık fotonunun yakalanma olasılığını artırır. Şu soru ortaya çıkıyor: Işık görsel pigmentler tarafından emildiğinde sinyaller nasıl ortaya çıkıyor?
Görsel pigmentler tarafından ışık emilimi
Işık çubuk pigmenti, rodopsin tarafından emildiğinde meydana gelen olaylar, psikofizyolojik, biyokimyasal ve moleküler teknikler kullanılarak incelenmiştir. Görsel pigment molekülü iki bileşenden oluşur: opsin adı verilen bir protein ve retina adı verilen bir kromofor, 11-cis-vitamin A-aldehit (Şekil 1). Kromoforun bileşiğe renk veren kimyasal bir grup içerdiği açıklığa kavuşturulmalıdır. Pigmentlerin absorpsiyon kapasitesinin nicel özellikleri, spektrofotometri kullanılarak incelenmiştir. Çubukların görsel pigmenti olan rodopsin, farklı dalga boylarında ışıkla aydınlatıldığında, yaklaşık 500 nm dalga boyuna sahip mavi-yeşil ışık en iyi şekilde emildi. Benzer bir sonuç, mikroskop altında tek bir çubuğun farklı dalga boylarında ışık huzmeleri ile aydınlatılmasıyla da elde edildi. Rodopsinin absorpsiyon spektrumu ile alacakaranlık ışığını algılamamız arasında ilginç bir ilişki bulunmuştur. İnsanlar üzerinde yapılan nicel psikofiziksel çalışmalar, yaklaşık 500 nm dalga boyuna sahip mavimsi yeşil gün ışığının karanlıkta alacakaranlık ışığının algılanması için optimal olduğunu göstermiştir. Çubukların aktif olmadığı ve sadece konilerin kullanıldığı gün boyunca, konilerin absorpsiyon spektrumuna karşılık gelen kırmızı renge en duyarlıyız (bundan daha sonra bahsedeceğiz).
Bir foton rodopsin tarafından absorbe edildiğinde, retinal fotoizomerizasyona uğrar ve 11-cis'ten trans konfigürasyonuna geçer. Bu geçiş çok hızlı gerçekleşir: yaklaşık 10-12 saniye içinde. Bundan sonra, pigmentin protein kısmı da bir dizi ara ürünün oluşumuyla birlikte bir dizi dönüşümsel değişikliğe uğrar. Protein parçasının konformasyonlarından biri olan metarhodopsin II, sinyal iletimi için en önemlisidir (bunu bu bölümde daha sonra tartışacağız). Şekil 2, aktif rodopsinin renginin giderilmesi ve rejenerasyonu sırasındaki olayların sırasını göstermektedir. 1 ms sonra Metarhodopsin II oluşur. Parçalanmasından sonra pigment rejenerasyonu birkaç dakika içinde yavaş yavaş gerçekleşir; bu, retinanın fotoreseptörlerden pigment epiteline taşınmasını gerektirir.
Rodopsin'in yapısı
Moleküler düzeyde, opsin proteini, her biri 20-25 amino asitten oluşan 7 hidrofobik bölge oluşturan ve 7 transmembran sarmal oluşturan 348 amino asit kalıntısından oluşur. Molekülün N-ucu hücre dışı boşlukta (yani çubuk diskin içinde) bulunur ve C-ucu sitoplazmada bulunur.
Şekil 1. Fotoreseptör zarına gömülü omurgalı rodopsin yapısı. Spiral, retinanın (siyah renkle gösterilen) yerini göstermek için biraz açılmıştır. C - C terminali, N - N terminali.
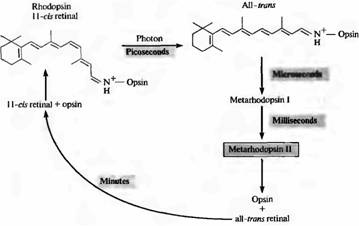
İncir. 2. Işıkta rodopsinin çiçeklenmesi. Karanlıkta, 11-cis-retinal, opsin proteinine sıkıca bağlıdır. Foton yakalama, retina tahtındaki tüm cis retinalin izomerizasyonuna yol açar. Bu durumda, opsin all-tron-retinal kompleksi hızla, opsin ve all-tron retinal'e ayrışan metarhodopsin II'ye dönüşür. Rodopsin rejenerasyonu, fotoreseptörler ve pigment hücrelerinin etkileşimine bağlıdır. Metarhodopsin II, ikinci haberci sistemini aktive eder ve sürdürür.
Retina, yedinci transmembran segmentinde bulunan bir lizin tortusu yoluyla opsine bağlanır. Opsin, adrenerjik ve muskarinik reseptörler gibi metabotropik aracı reseptörleri de içeren 7 transmembran alanına sahip bir protein ailesine aittir. Rodopsin gibi, bu reseptörler G-protein aktivasyonu yoluyla ikinci habercilere sinyal verir. Rodopsin karanlıkta oldukça kararlıdır. Bayor, rodopsin molekülünün kendiliğinden termal izomerizasyonunun yaklaşık 3000 yıl veya fotoizomerizasyondan 1023 daha fazla sürdüğünü hesapladı.
Koniler ve renkli görme
19. yüzyılda Young ve Helmholtz tarafından yürütülen şaşırtıcı araştırma ve deneyler, önemli konu renk görme hakkında ve bilim adamlarının kendileri bu fenomenin net ve doğru bir açıklamasını yaptılar. Üçünün varlığına dair vardıkları sonuç çeşitli tipler renkli fotoreseptörler zamana direnmiş ve daha sonra moleküler düzeyde doğrulanmıştır. Yine ışık ve ses, renk ve ses tonu algısını karşılaştıran Helmholtz'dan alıntı yapabiliriz. Özellikle 19. yüzyılda yaygın olan kafa karıştırıcı vitalistik kavramlarla karşılaştırıldığında, düşüncesinin netliği, gücü ve güzelliğine imrenilebilir:
Renk tonlarındaki tüm farklılıklar, üç ana rengin çeşitli oranlardaki kombinasyonuna bağlıdır ... kırmızı, yeşil ve menekşe ... Tıpkı güneş ışığının ve sıcaklığının algılanmasının ... güneş ışınlarının vurup vurmadığına bağlı olduğu gibi ... sinirler, görme alıcılarından veya termal duyarlılık alıcılarından gelir. Young'ın hipotezinde öne sürdüğü gibi, farklı renklerin algılanmasındaki farklılık, bu ışık tarafından 3 tip fotoreseptörden hangisinin daha fazla aktive edildiğine bağlıdır. Her üç tür de olduğunda eşit olarak heyecanlı, beyaza dönüyor ...
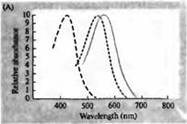
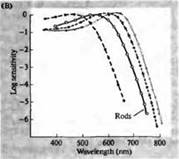
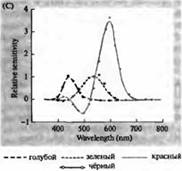
Pirinç. 3. İnsan fotoreseptörlerinin duyarlılık spektrumları ve çeşitli görsel pigmentler. (A) Camgöbeği, yeşil ve kırmızıya karşılık gelen dalga boylarında absorpsiyon zirvelerini gösteren üç renkli görsel pigmentin duyarlılık eğrileri. (B) Makaklarda konilerin mavi, yeşil ve kırmızıya ve çubukların (siyah renkle gösterilmiştir) duyarlılık spektrumları. Yanıtlar, emme elektrotları kullanılarak kaydedildi, ortalaması alındı ve normalleştirildi. Çubuk spektrumunun eğrileri, insanlarda görsel pigmentlerin çalışmasında elde edildi. (C) Bir renk duyarlılığı testi kullanılarak maymun ve insan koni spektrumlarının karşılaştırılması. Sürekli eğri, farklı dalga boylarında ışık ile sunulduğunda insanlarda renge duyarlılığı belirlemek için bir deneyi gösterir. Noktalı çizgi, dış segmente giden yolda lens ve pigmentlerdeki ışığın absorpsiyonunu düzelttikten sonra, ayrı konilerdeki akımların kaydına dayalı olarak tahmin edilen sonuçları gösterir. Her iki deneyin sonuçları arasındaki uyum şaşırtıcı derecede yüksektir.
Beyaz bir ekrana aynı anda farklı renklerde iki ışık huzmesi yansıtırsak... sadece bir renk görürüz, bu iki renkten aşağı yukarı farklı. Gözü kuru bir göz ile karşılaştırırsak, üç ana rengin karışımı ile dış ışığın bileşimindeki tüm tonları algılayabildiğimiz olağanüstü gerçeğini daha iyi anlayabiliriz ... Ses durumunda ... biz daha uzun dalgaları alçak tonlar ve kısa dalgalar olarak duyun - yüksek ve delici olarak, ayrıca kulak aynı anda birçok ses dalgasını yakalayabilir, yani. birçok not. Ancak, bu durumda β tek bir karmaşık akorda birleşmezler, tıpkı farklı renklerin ... tek bir karmaşık renkte birleşmesi gibi. Değiştirirsek göz farkı söyleyemez turuncu renk kırmızıya veya sarıya; ama notaların do ve mi sesini aynı anda duyarsak, böyle bir ses bize nota re gibi gelmiyor. Kulak, müzik tonlarını göz renkleri algıladığı gibi algılıyorsa, her akor, biri çok düşük, biri çok yüksek ve biri orta olmak üzere üç sabit notanın bir kombinasyonu ile temsil edilebilir ve tüm olası müzik efektlerini yalnızca bu notaların göreli ses yüksekliğini değiştirerek üretebilir. üç nota. .. Ancak sonsuz sayıda gölge ve geçiş sayesinde renklerin birinden diğerine yumuşak geçişini görebiliriz... Renklerin her birini algılama şeklimiz... gergin sistem. Şu anda kabul edilmelidir ki, ne insanlarda ne de tetrapodlarda renk algısı teorisini doğrulamak için anatomik bir temel tanımlanmamıştır.
Bu doğru ve ileri görüşlü tahminler, bir dizi farklı gözlemle doğrulanmıştır. Spektrofotometri kullanarak, Wald, Brouck, McNicol ve Dartnell ve arkadaşları, insan retinasında farklı pigmentlere sahip üç tip koninin varlığını gösterdi. Baylor ve meslektaşları ayrıca maymunların ve insanların konilerinden gelen akımları yönlendirmeyi başardılar. Üç koni popülasyonunun, spektrumun mavi, yeşil ve kırmızı kısımlarına karşı farklı fakat örtüşen hassasiyet aralıklarına sahip olduğu bulundu. Elektrik sinyallerinin uyarılması için optimal dalga boyları, gözün renk spektrumuna duyarlılığını ölçmek için spektrofotometri ve psikofiziksel deneyler kullanılarak oluşturulan görsel pigmentler tarafından ışık absorpsiyonunun zirveleriyle tam olarak çakıştı. Sonunda Natais, opsin pigmentini kodlayan genleri kırmızı, yeşil ve maviye duyarlı üç tip konide klonladı ve sıraladı.
O halde, çeşitli görsel pigmentlerin molekülleri, tercihli olarak belirli bir dalga boyundaki ışığı nasıl yakalayabilir? Rodopsin'in çubukların görsel pigmenti olduğu ve konilerin üç görsel pigmentinin de aynı kromoforu, 11-cis-retinal'i içerdiği ortaya çıktı. Ancak pigmentin protein kısmının amino asit dizileri birbirinden farklıdır. Sadece birkaç amino asitteki farklılıklar, onların spektruma karşı farklı hassasiyetlerini açıklar.
renk körlüğü
Bir tür fotoreseptör kendi başına rengi algılayamazken, Şekil l'de gösterildiği gibi üç tip koni. 4, zaten yetenekli.
Prensipte, renk tanıma için farklı pigmentlere sahip iki tip koni yeterli olacaktır, ancak bu durumda dalga boylarının bir dizi kombinasyonu eşit olarak algılanacaktır. Benzer bir durum, bir kişi renk körlüğünden muzdarip olduğunda ortaya çıkar. Nathans tarafından gösterildiği gibi, bu tür insanlar, pigmentlerden birinin yokluğuna yol açan genetik kusurlara sahiptir. Bilimin mevcut durumunun bakış açısından, moleküler mekanizmaların Young ve Helmholtz'un parlak ve şaşırtıcı derecede doğru düşüncelerini ne kadar güzel bir şekilde doğruladığına şaşırmadan edemiyoruz.
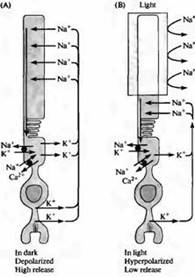
Pirinç. 4. Bir çubukta "karanlık" akım. (A) Karanlıkta, sodyum iyonları çubukların dış segmentindeki katyon kanallarından geçerek depolarizasyona neden olur; kalsiyum iyonları da bu katyon kanallarından geçebilir. Potasyum akımının zarın iç bölümünde dışa doğru akması nedeniyle akım döngüsü çubuğun kıstağından geçer. (B) Dış segment aydınlatıldığında, cGMP'nin hücre içi konsantrasyonundaki azalma nedeniyle kanallar kapanır ve çubuk hiperpolarize olur. Hiperpolarizasyon, nörotransmitterin salınımında bir azalmaya yol açar. Sodyum, potasyum ve kalsiyum konsantrasyonları, çubuğun içinde özel pompalar ve iç segment bölgesinde (siyah daireler) bulunan iyon değiştiriciler tarafından korunur. Kalsiyum taşıyıcıları da dış segmentte bulunur.
Renk görme ve renk körlüğünün temel özelliklerinin fotoreseptörlerin kendisinde bulunması gerektiği fikri, doğrudan fizyolojik ölçümlerle ve ayrıca pigmentlerin yapısındaki genetik ve protein seviyelerindeki farklılıkların incelenmesiyle doğrulandı. -40 mV, denge potasyum potansiyelinden uzak olan E K, yani - 80 mV.

Pirinç. 5. Çubukların dış segmentindeki sodyum kanallarının açılmasında cGMP'nin rolü. Tek kanalların aktivitesi, farklı konsantrasyonlarda cGMP ile çözelti içinde olan bir içten dışa yama kelepçesi kullanılarak kaydedildi. Kanalın açılması akımın yukarı doğru sapmasına neden olur. Kontrolde kanal açma frekansı çok düşüktü, cGMP'nin eklenmesi, doğrudan cGMP konsantrasyonuna bağlı olan tek kanal açma frekansında bir artışa yol açtı.
Karanlıkta gelen akım, dış segmentin katyon kanalları boyunca bir elektrokimyasal gradyanı takiben esas olarak sodyum iyonları tarafından taşınır. Işığın etkisi altında fotoreseptörün hiperpolarizasyonu, bu kanalların kapanmasından kaynaklanır, bu da membran potansiyelinde E K'ye doğru bir kaymaya yol açar.
Fotoreseptör kanallarının özellikleri
Fizyolojik koşullar altında dış bölümün katyon kanalları, 12.5:1.0:0.7'lik bir kalsiyum/sodyum/potasyum iyonu iletkenlik oranına ve yaklaşık 0.1 pS 21'lik bir tek kanal iletkenliğine sahiptir. Sodyum konsantrasyonu, kalsiyum konsantrasyonundan çok daha yüksek olduğu için, gelen akımın yaklaşık %85'i sodyum iyonlarıdır. Potasyum iyonlarının itici gücü hücrenin dışına yönlendirilir. Kalsiyum iyonları kanaldan geçtiğinde, bu iyonlar gözenek duvarına sıkıca bağlanır ve böylece diğer iyonların iletimini engeller. Bu nedenle, hücre dışı ortamdan kalsiyumun çıkarılması, iletkenliği 25 pS'ye yükselen kanallardan potasyum ve sodyum iyonlarının daha kolay geçişine yol açar.
Fesenko, Yau, Baylor, Strayer ve meslektaşları, döngüsel HMP'nin diskten membran yüzeyine dahili bir sinyal taşıyıcı rolü oynadığını gösterdi. Şekil 4'te gösterildiği gibi, sitoplazmada yüksek bir cGMP konsantrasyonu katyon kanallarını açık tutar. Membranın iç yüzeyinden cGMP konsantrasyonunda bir azalma ile katyon kanallarının açılması nadir görülen bir olay haline gelir. Böylece, fotoreseptörlerin zar potansiyeli sitoplazmadaki cGMP konsantrasyonunun bir yansımasıdır: cGMP konsantrasyonu ne kadar yüksek olursa, hücrenin depolarizasyonu o kadar büyük olur. cGMP konsantrasyonu da gelen ışığın yoğunluğuna bağlıdır. Işık yoğunluğundaki bir artış, cGMP konsantrasyonunda bir azalmaya yol açar ve açık kanalların oranını azaltır. cGMP'nin yokluğunda, hemen hemen tüm kanallar kapalıdır ve dış segment zarının direnci, lipid çift tabakasının direncine yaklaşır.
cGMP kapılı kanalların moleküler yapısı
Çubukların dış bölümünün iyon kanalları için cDNA izole edildi ve bu kanalların alt birimlerinin insan, sığır, fare ve tavuk retinalarındaki amino asit dizileri belirlendi. Bunlar ve diğer cGMP kapılı iyon kanalları için - örneğin koku alma sisteminde bulunanlar gibi - önemli DNA benzerliği bulunmuştur.
Fotoreseptörlerde sinyal iletimi
Rodopsinin fotoizomerizasyonu zar potansiyelinde nasıl bir değişikliğe yol açar? Çubuk ve konilerdeki elektrik sinyallerini uyarmak için bir çeşit dahili aracıya ihtiyaç olduğu yıllar içinde anlaşılmıştır. Çubuğun dış segmenti bölgesindeki fotonların absorpsiyonuyla ilgili bilgilerin bir taşıyıcı tarafından iletildiğinden şüphelenmenin nedenlerinden biri, pigment rodopsinin kendisinin diskin içinde yer alması ve sinyalin sitoplazma yoluyla sitoplazma boyunca yayılmasıydı. dış membran. İkinci neden, yanıtta önemli bir artış oldu. Kaplumbağalarda fotoreseptörler üzerinde çalışan Baylor ve meslektaşları, sadece bir foton emildiğinde ve 108 pigment molekülünden biri aktive edildiğinde, zar iletkenliğinde ve kaydedilen elektrik sinyallerinde bir azalmanın meydana geldiğini gösterdi.
Aktive edilmiş bir fotopigment molekülünün membran potansiyelini değiştirdiği olayların sırası, iki elektrotlu potansiyel kenetleme ve moleküler biyoloji teknikleri kullanılarak çubukların ve konilerin dış bölümleri incelenerek açıklanmıştır. Bir ışık fotonunun absorpsiyonundan bir elektrik sinyaline sinyal iletimi şeması, Şek. 3.
Karanlıkta, çubukların ve konilerin dış bölümlerinde sürekli bir gelen "karanlık" akım akar. Sonuç onların membran sistemidir. Bu kanalların zar bölgeleri, özellikle S4 bölgesi ve iyon porunu oluşturan bölgede diğer katyon seçici kanallarla yapısal benzerlikler göstermektedir. Fotoreseptör iyon kanalları, moleküler ağırlığı sırasıyla 63 ve 240 kDa olan en az 2 farklı protein alt biriminden oluşan tetramerlerdir.
Hücre içi nükleotid bağlanma bölgeleri, alt birimlerin karboksil ucunun yakınında bulunur. Bu alt birimlerin oositlerde ekspresyonu, çubukların dış segmentlerinde bulunan kanallara benzer özelliklere sahip katyon kanallarının oluşumuna yol açar: cGMP tarafından aktive edilirler ve beklenen iletkenlik ve geçirgenlik oranlarına sahiptirler.

Pirinç. 6. Fotopigment molekülünün aktivasyonu üzerine G-protein aktivasyonunun mekanizması. G-protein transdusin, metarhodopsin II varlığında GTP'ye bağlanır, bu da fosfodiesterazın aktivasyonuna yol açar ve bu da cGMP'yi hidrolize eder. cGMP konsantrasyonu düştüğünde, sodyum kanalları kapanır.
Döngüsel GMP'nin metabolik kaskadı
cGMP konsantrasyonunda bir azalmaya ve ardından iyon kanallarının kapanmasına yol açan olaylar zinciri Şekil 5'te gösterilmektedir. cGMP'nin hücre içi konsantrasyonundaki bir azalmaya ışık neden olur ve fotopigmentin parçalanmasında bir ara ürün olan metarhodopsin II'nin oluşumuna yol açar. Metarhodopsin II, sırayla, 3 polipeptit zincirinden oluşan G-protein transdusin üzerinde etki eder.
Metarhodopsin II ve transdusinin etkileşimi, GDP'nin G-proteinine bağlı molekülünün GTP ile değiştirilmesine yol açar. Bu, alt birimlerden ayrılan G-protein alt birimini aktive eder ve sırayla membran bölgesinde bulunan fosfodiesterazı aktive eder: cGMP'yi hidrolize eden bir enzim. cGMP konsantrasyonu düşer, daha az açık iyon kanalı vardır, çubuk hiperpolarize olur. Kaskad, aktif metarhodopsin II'nin C-terminalinin fosforilasyonu ile kesintiye uğrar. cGMP'nin katyon kanallarının durumunu düzenlemedeki kilit rolü, biyokimyasal deneylerle doğrulanmıştır. Fotoreseptörlerin aydınlatılması, hücredeki cGMP seviyesinde %20'lik bir düşüşe neden olabilir.
Işığa maruz kaldığında depolarize olan omurgalı reseptörleri
Yukarıda verilen foto-alma mekanizmasının ilginç bir istisnası, bazı omurgalı reseptörleridir. Kertenkelelerin başlarının üstünde bulunan üçüncü bir gözü vardır. Ana (yan) gözler tarafından algılanana benzer bir görüntüyü algılayabilen küçük "koniler" içerir. Bununla birlikte, bu fotoreseptörler, aydınlatma üzerine depolarize olmaları bakımından dikkate değerdir. Buradaki nükleotid kapılı kanallar, bir istisna dışında diğer omurgalıların fotoreseptörlerine benzer bir yapıya ve işleve sahiptir: fotoreseptör ve G-proteininin aktivasyonu, cGMP konsantrasyonunda bir artışa yol açar. Sonuç olarak, dış segmentin kanalları açılır ve katyonlar hücreye yönlendirilerek "hafif" bir akım oluşturur. Bunun nedeni karanlıkta fosfodiesteraz aktivitesinin inhibisyonudur. Sonuç olarak, kertenkele gözünde aşağıdaki olaylar dizisi meydana gelir: ışık - [cGMP] - dış segmentin katyon kanallarının açılması - depolarizasyon.
cGMP kademesinde sinyal amplifikasyonu
İki aşamalı cGMP kaskadı, çubukların ışığa karşı olağanüstü hassasiyetini açıklayan orijinal sinyalin önemli bir amplifikasyonunu sağlar. İlk olarak, bir aktif metarhodopsin II molekülü, GDP yerine birçok GTP molekülünün eklenmesini katalize eder ve böylece yüzlerce G-protein alt birimini serbest bırakır. İkincisi, her bir alt birim, diskte sitoplazmada bulunan çok sayıda cGMP molekülünü parçalayabilen ve böylece çok sayıda kanalın kapanmasına yol açabilen bir fosfodiesteraz molekülünü aktive eder.
Tek ışık kuantumuna yanıt olarak sinyaller
Tek ışık kuantumunun algılanan ışık hissine neden olabileceği veriler çok sayıda sorular. Bu tek cevap ne kadar büyük? Bu sinyal gürültü tabanından nasıl ayrılıyor? Ve bu tür bilgiler retinadan daha yüksek görme merkezlerine nasıl güvenilir bir şekilde iletilir? Baylor ve meslektaşları, tek ışık kuantumuna yanıt olarak sinyalleri ölçmek için kurbağaların, maymunların ve insanların retinalarındaki ayrı çubuklardan gelen akımları kaydettiler. Bu deneyler, zayıf ışık parlamalarının algılanması gibi karmaşık bir sürecin, tek tek moleküller düzeyinde meydana gelen değişikliklerle nasıl ilişkilendirilebileceğini gösteren benzersiz bir deney örneğidir.
Bu deneyler için bir hayvan veya kadavra materyalinden bir retina parçasının izole edilmesi prosedürü karanlıkta gerçekleştirilmelidir. Akımı ölçmek için çubuğun dış kısmı ince bir pipete emilir. Beklendiği gibi, bu deneyler karanlıkta akımın sürekli olarak dış segmentin içinde aktığını göstermektedir. Işık yanıp sönmeleri, dış segmentteki kanalları kapatarak "karanlık" akımında bir azalmaya neden olur. Akım genliği küçüktür ve soğurulan fotonların sayısıyla orantılıdır. Bazen flaş tek bir tepkiye, bazen çift bir tepkiye ve bazen de hiç tepkiye neden olmaz.
Maymun çubuklarında, bir fotonun absorpsiyonuna yanıt olarak akımdaki azalma yaklaşık 0,5 pA'dır. Bu, yaklaşık 300 kanalın kapanmasına karşılık gelir, yani. tüm kanalların %3 ila %5'i karanlıkta açılır. Bu, metabolik sinyaldeki önemli bir artış nedeniyle elde edilir. cGMP çağlayanı. Ayrıca, daha önce bahsedilen görsel pigmentlerin aşırı stabilitesi nedeniyle, rastgele izomerizasyon ve yanlış kanal kapanması çok nadir görülen olaylardır. Bu, tek tek ışık kuantumlarının etkilerinin çok düşük DC parazitli bir arka planda öne çıkmasına neden olur. Fotoreseptörler arasındaki boşluk bağlantısı elektrik kuplajının, arka plan gürültüsünü azaltan ve çubuk tepkilerinin tek kuantaya sinyal-gürültü oranını iyileştiren ek bir yumuşatma etkisi sağladığı gösterilmiştir.
Edebiyat
1. Finn, J.T., Grunwald, M.E ve Yau, K-W. 1996. Döngüsel nükleotid kapılı iyon kanalları: Farklı işlevlere sahip genişletilmiş bir aile. Annu. Rev. Physiol.58: 395-426.
2. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y, Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S. ve Okada, M. 1998. Glutamat reseptörleri : Beyin fonksiyonu ve sinyal iletimi.
GÖRSEL PİGMENTLER(lat. pigmentum boya) - retinanın fotoreseptörlerinin ışığa duyarlı pigmentleri. 3. Bir ışık darbesinin enerjisini algılayan maddeler karmaşık bir fotokimyasal döngüye girer. 3 p (bir koni veya bir çubuk) içeren retinanın ayrı bir görsel reseptörünün uyarılmış bir duruma geçtiği ve alınan bilgiyi optik sinir boyunca c'ye ilettiği dönüşümler. n. itibaren. Bir gözün retinasının görsel hücrelerinin fotoreseptör membranının ana yapısal ve fonksiyonel parçası olan 3. maddeler bu nedenle görme mekanizmalarında kilit bir rol oynar (bkz.).
Görsel pigmentlerin isimlendirilmesi ve yapısı. Omurgalıların ve omurgasızların üzerinde çalışılan tümü, suda çözünmeyen membran proteini opsin ve onunla bağlantılı kromoforun (retinal) kompleksleridir. Retina veya A vitamini aldehit, retinal1 ve retinal2 olmak üzere iki şekilde bulunabilir.
Bir kromofor 3'ün doğası gereği, madde iki sınıfa ayrılır - retinal1 içeren rodopsinler (bkz.) ve retinal2 içeren porfiropsinler. Rodopsinler tüm kara ve deniz hayvanlarının gözünün retinasında bulunurken, porfiropsinler tatlı su hayvanlarının retinasında bulunur. Bazı balıklar ve amfibiler aynı anda 3 p retina içerir! ve retinal. Retinanın çubuklarına veya konilerine özgü opsinlerdeki farklılıklar temelinde 3. s.'yi sınıflandırmaya yönelik girişimler vardır. Örneğin, rodopsin, çubuk opsinli bir retinal1 kompleksidir, iyodopsin, koni opsinli bir retinal1 kompleksidir, porfiropsin, çubuk opsinli retinal2'dir, retinal-koni opsin kompleksi siyanopsini oluşturur. Ancak en az beş farklı opsin olduğu için 3. maddeyi opsin bazında sınıflandırmak son derece zordur.
Bilinen tüm 3. maddelerden boğa, kurbağa ve kalamarın gözlerinden izole edilen rodopsinler en kapsamlı şekilde araştırılmıştır. Onların savunması. 30-40 bin mertebesinde ağırlık (kütle), her molekül yakl. 400 amino asit ve bir kromofor. Ek olarak, oligosakkarit zinciri 3. p.: 3 glukozamin radikali, 2 mannoz, 1 galaktoz molekülüne dahildir. Lipitler (ch. arr. fosfolipidler) molekül 3 ile güçlü bir kompleks oluştururlar. s. 3. Temel spektral özelliklerini korurken (bkz.
saf retina vardır Sarı, absorpsiyon spektrumunun maksimumu 370 nm bölgesindedir. Opsin renksizdir, maksimum absorpsiyon ultraviyole bölgesindedir (yaklaşık 280 nm). Rodopsin molekülünün rengi kırmızımsı pembedir, maksimum absorpsiyon spektrumu yakl. 500 nm. Kompleksin oluşumu sırasında bu kadar güçlü bir spektral kaymanın nedeni (370'den 500 nm'ye - sözde batokromik kayma) henüz kesin bir açıklama almadı.
Rodopsinlerin ve porfiropsinlerin absorpsiyon spektrumlarının maksimumu, görünür spektrumun oldukça geniş bir bölgesini yakalar - rodopsinler için 433 ila 562 nm ve porfiropsinler için 510 ila 543 nm. Porfiropsinler ayrıca bir kurbağa, sazan ve tatlı su kaplumbağasının iribaşlarının 3.p konilerini, yani 620 nm'de maksimum absorpsiyon spektrumuna sahip siyanopsin'i içeriyorsa, bu bölge daha da geniştir. Mikrospektrofotometri yöntemlerinin geliştirilmesi, hayvanlarda ve insanlarda birçok tekli fotoreseptör hücre tipinin absorpsiyon spektrumlarının belirlenmesini mümkün kılmıştır. Elde edilen verilere göre, insan retinasının 3. p.'si aşağıdaki absorpsiyon spektrum maksimumlarına sahiptir: çubuklar 498, mavi-, yeşil- ve kırmızıya duyarlı koniler - sırasıyla 440, 535 ve 575 nm.
3. s.'nin çalışması Almanca olarak başladı. 1851'de bir kurbağanın gözünden alınan pembemsi-mor retinanın nasıl ışıkta önce sarımsı, sonra beyazımsı olduğunu anlatan araştırmacı H. Muller. 1877'de F. Boll, retinanın görme hücrelerinde bir tür kırmızı ışığa duyarlı maddenin bulunduğu ve bu maddenin renginin bozulmasının görme mekanizması ile ilişkili olduğu sonucuna vararak bu fenomeni de tanımladı. 3. s. çalışmasında büyük bir değer Kuhne'ye aittir (W. Kuhne, 1877), Krom 3. s.'yi izole etmeyi ve ayrıntılı olarak incelemeyi başardı. Çıkardığı 3. s.'yi görsel mor olarak adlandırdı, protein yapısını belirledi, bazı spektral özelliklerini ve foto dönüşümlerini araştırdı, 3. s.'nin karanlıkta iyileşme yeteneğini keşfetti. Amer., 3. s çalışmasına büyük katkı sağlamıştır. biyokimyacı ve fizyolog J. Wald.
Görsel pigmentlerin fototransformasyonları. Işığın 3. s. üzerindeki etkisi altında, içlerinde bir fotokimyasal döngü meydana gelir. retinanın cis-trans izomerizasyonunun birincil fotokimyasal reaksiyonuna dayanan dönüşümler (bkz. İzomerizm). Bu durumda kromofor ile protein arasındaki bağ bozulur. 3. s.'nin dönüşüm dizisi şu şekilde temsil edilebilir: rodopsin (kromofor cis formundadır) -> prelumirhodopsin -> lumirhodopsin -> metarhodopsin I -> metarhodopsin II -> opsin proteini -> trans formda kromofor . Enzim - retinol dehidrojenazın etkisi altında - ikincisi, çubukların ve konilerin dış bölümlerinden retinanın pigment tabakasının hücrelerine gelen A vitaminine geçer. Gözü karartırken, rejenerasyon meydana gelir 3. s., bir kesimin uygulanması için, bir kromofor (A vitamini aldehit) oluşumu için başlangıç ürünü olarak hizmet eden bir A vitamini cis-izomerinin varlığı gereklidir. . Vücutta A vitamini eksikliği veya yokluğu ile, rodopsin oluşumu sözde alacakaranlık görme bozukluğunun bir sonucu olarak bozulabilir ve gelişebilir. gece körlüğü (bkz. Hemeralopi). Rodopsinin fototransformasyon sürecinde, reseptör hücrede lumirhodopsinin metarhodopsin I'e geçiş aşamasında, sözde parlak bir flaşa yanıt olarak meydana gelir. erken (kısa gecikmeli) reseptör potansiyeli. Ancak, o değil görsel sinyal fotoreseptör membranında 3. p.'nin dönüşüm mekanizmasını incelemek için testlerden biri olarak hizmet edebilmesine rağmen. İşlevsel değer sözde. geç reseptör potansiyeli, rogo'ya kadar olan latent dönem (5-10 msn), metarhodopsin II'nin oluşum zamanı ile orantılıdır. Metarhodopsin I'in metarhodopsin II'ye geçişinin reaksiyonunun görsel bir sinyalin ortaya çıkmasını sağladığı varsayılmaktadır.
3. öğeler ışıkta sürekli renk değiştirdiğinden, sürekli restorasyonu için mekanizmalar olmalıdır. Bazıları son derece hızlıdır (fotorejenerasyon), diğerleri oldukça hızlıdır (biyokimyasal, rejenerasyon, karanlık), diğerleri yavaştır (3. s sentezi. Görsel hücredeki fotoreseptör zarının sürekli yenilenmesi sırasında). Fotorejenerasyonun omurgasız hayvanlarda değeri olan fiziol vardır (örneğin, kafadanbacaklılar- Kalamar ahtapotu). Biyokimya mekanizmasında. rejenerasyon 3. s. omurgalılarda, görünüşe göre, trans-retinalin (veya trans-vitamin A) tekrar cis-izomer formuna izomerizasyonunu sağlayan izomeraz enzimi (bakınız) tarafından önemli bir rol oynar. Ancak böyle bir enzimin varlığına dair kesin bir kanıt yoktur. Sistemde retinal ve opsin'in 11-cis-izomeri varlığında bir molekül 3. s. oluşumunun aynı reaksiyonu, enerji harcaması olmadan kolayca gerçekleşir. Ağartılmış rodopsinin fosforilasyon reaksiyonuna yeteneği bulunur (bakınız); bu reaksiyonun, görsel hücrenin ışık adaptasyonu mekanizmasındaki bağlantılardan biri olduğu varsayılmaktadır.
Kaynakça: Ackerman Yu Biyofizik, çev. İngilizce'den, M., 1964; Willie K. ve Det e V. Biology, çev. İngilizceden, M., 1974, bibliyografya; Konev S. V. ve Volotovsky I. D. Moleküler fotobiyolojiye giriş, s. 61, Minsk, 1971; Ostrovsky M. A. ve Fedorovich I. B. Fotoreseptör zarında fotoindüklenen değişiklikler, kitapta: Biol'ün yapısı ve işlevleri, zarlar, ed. A.S. Troshina ve diğerleri, s. 224, M., 1975, bibliogr.; Duyusal sistemlerin fizyolojisi, ed. G. V. Gershuni, bölüm 1, s. 88, L., 1971; Görsel pigmentlerin biyokimyası ve fizyolojisi, ed. H. Langer, B.a. o., 1973; Duyusal fizyoloji el kitabı, ed. H.A.R. Jung a. o., v. 7, pt 1-2, B., 1972.
M.A. Ostrovsky.


