vizuelni pigmenti. Rudimentarna zona epiderme kože sastoji se od slojeva. Dnevni i noćni vid
Vizualni pigmenti koncentrirani su u membranama vanjskih segmenata. Svaki štapić sadrži oko 10 8 molekula pigmenta. Oni su organizirani u nekoliko stotina diskretnih diskova (oko 750 u majmunskom štapiću) koji nisu povezani s vanjskom membranom. U čunjićima pigment se nalazi u posebnim pigmentnim naborima koji su nastavak vanjske stanične membrane fotoreceptora. Molekuli pigmenta čine oko 80% svih proteina diska. Vizualni pigmenti su tako gusto upakovani u membrane vanjskog segmenta da razmak između dva molekula vizualnog pigmenta u štapiću ne prelazi 10 nm. Ovako gusto pakovanje povećava vjerovatnoću da će foton svjetlosti koji prolazi kroz sloj fotoreceptorskih ćelija biti uhvaćen. Postavlja se sljedeće pitanje: kako nastaju signali kada svjetlo apsorbiraju vizualni pigmenti?
Apsorpcija svjetlosti vizualnim pigmentimaDogađaji koji se dešavaju kada svetlost apsorbuje štap pigment, rodopsin, proučavani su psihofiziološkim, biohemijskim i molekularnim tehnikama. Molekul vizualnog pigmenta sastoji se od dvije komponente: proteina, nazvanog opsin, i hromofora, 11-cis-vitamin A-aldehida, koji se zove retinal (slika 1). Treba pojasniti da hromofor sadrži hemijsku grupu koja daje boju spoju. Kvantitativne karakteristike apsorpcionog kapaciteta pigmenata proučavane su spektrofotometrijom. Kada je rodopsin, vizuelni pigment štapića, bio osvetljen svetlošću različitih talasnih dužina, najbolje se apsorbovala plavo-zelena svetlost talasne dužine od oko 500 nm. Sličan rezultat je također dobiven osvjetljavanjem jednog štapa pod mikroskopom snopovima svjetlosti različitih talasnih dužina. Otkriveno je zanimljiva zavisnost između spektra apsorpcije rodopsina i naše percepcije sumračne svjetlosti. Kvantitativne psihofizičke studije provedene na ljudima pokazale su da je plavkasto-zelena dnevna svjetlost s talasnom dužinom od oko 500 nm optimalna za percepciju sumračnog svjetla u mraku. Tokom dana, kada su štapići neaktivni i koriste se samo čunjevi, najosjetljiviji smo na crvenu boju koja odgovara spektru apsorpcije čunjeva (o tome ćemo kasnije).
Kada jedan foton apsorbuje rodopsin, retinal prolazi kroz fotoizomerizaciju i prelazi iz 11-cis u trans konfiguraciju. Ovaj prelaz se dešava veoma brzo: za oko 10-12 sekundi. Nakon toga, proteinski dio pigmenta također prolazi kroz niz transformacijskih promjena, uz formiranje niza međuprodukata. Jedna od konformacija proteinskog dijela, metarodopsin II, najvažnija je za transdukciju signala (o tome ćemo raspravljati kasnije u ovom poglavlju). Slika 2 prikazuje redoslijed događaja tokom dekolorizacije i regeneracije aktivnog rodopsina. Metahodopsin II se formira nakon 1 ms. Regeneracija pigmenta nakon njegovog raspada se odvija polako, u roku od nekoliko minuta; ovo zahtijeva transport retine od fotoreceptora do pigmentnog epitela.
Struktura rodopsinaNa molekularnom nivou, opsin protein se sastoji od 348 aminokiselinskih ostataka, formirajući 7 hidrofobnih zona, od kojih se svaka sastoji od 20-25 aminokiselina, koje čine 7 transmembranskih spirala. N-terminus molekula nalazi se u ekstracelularnom prostoru (tj. unutar diska štapa), a C-terminus se nalazi u citoplazmi.
U mraku, 11-cis-retinal je čvrsto vezan za protein opsin. Hvatanje fotona dovodi do izomerizacije svih cis retinala u retinalnom tronu. U ovom slučaju, opsin all-tron-retinal kompleks brzo se pretvara u metarodopsin II, koji se disocira u opsin i all-tron retinal. Regeneracija rodopsina zavisi od interakcije fotoreceptora i pigmentnih ćelija. Metarhodopsin II aktivira i održava sistem drugog glasnika.
Retinal je povezan sa opsinom preko ostatka lizina koji se nalazi u sedmom transmembranskom segmentu. Opsin pripada porodici proteina sa 7 transmembranskih domena, što uključuje i metabotropne medijatorne receptore, kao što su adrenergički i muskarinski receptori. Poput rodopsina, ovi receptori signaliziraju sekundarnim glasnicima putem aktivacije G-proteina. Rodopsin je izuzetno stabilan u mraku. Bayor je izračunao da spontana termička izomerizacija molekula rodopsina traje oko 3000 godina, ili 10 23 više nego za fotoizomerizaciju.
vizuelni pigmenti
vizuelni pigmenti
Struktura rodopsina
Konusi i vid boja
daltonizam
Književnost
vizuelni pigmenti
Vizualni pigmenti koncentrirani su u membranama vanjskih segmenata. Svaki štapić sadrži oko 10 8 molekula pigmenta. Oni su organizirani u nekoliko stotina diskretnih diskova (oko 750 u majmunskom štapiću) koji nisu povezani s vanjskom membranom. U čunjićima pigment se nalazi u posebnim pigmentnim naborima koji su nastavak vanjske stanične membrane fotoreceptora. Molekuli pigmenta čine oko 80% svih proteina diska. Vizualni pigmenti su tako gusto upakovani u membrane vanjskog segmenta da razmak između dva molekula vizualnog pigmenta u štapiću ne prelazi 10 nm. Ovako gusto pakovanje povećava vjerovatnoću da će foton svjetlosti koji prolazi kroz sloj fotoreceptorskih ćelija biti uhvaćen. Postavlja se sljedeće pitanje: kako nastaju signali kada svjetlo apsorbiraju vizualni pigmenti?
Apsorpcija svjetlosti vizualnim pigmentima
Događaji koji se dešavaju kada svetlost apsorbuje štap pigment, rodopsin, proučavani su psihofiziološkim, biohemijskim i molekularnim tehnikama. Molekul vizualnog pigmenta sastoji se od dvije komponente: proteina, nazvanog opsin, i hromofora, 11-cis-vitamin A-aldehida, koji se zove retinal (slika 1). Treba pojasniti da hromofor sadrži hemijsku grupu koja daje boju spoju. Kvantitativne karakteristike apsorpcionog kapaciteta pigmenata proučavane su spektrofotometrijom. Kada je rodopsin, vizuelni pigment štapića, bio osvetljen svetlošću različitih talasnih dužina, najbolje se apsorbovala plavo-zelena svetlost talasne dužine od oko 500 nm. Sličan rezultat je također dobiven osvjetljavanjem jednog štapa pod mikroskopom snopovima svjetlosti različitih talasnih dužina. Pronađena je zanimljiva veza između spektra apsorpcije rodopsina i naše percepcije sumračne svjetlosti. Kvantitativne psihofizičke studije provedene na ljudima pokazale su da je plavkasto-zelena dnevna svjetlost s talasnom dužinom od oko 500 nm optimalna za percepciju sumračnog svjetla u mraku. Tokom dana, kada su štapići neaktivni i koriste se samo čunjevi, najosjetljiviji smo na crvenu boju koja odgovara spektru apsorpcije čunjeva (o tome ćemo kasnije).
Kada jedan foton apsorbuje rodopsin, retinal prolazi kroz fotoizomerizaciju i prelazi iz 11-cis u trans konfiguraciju. Ovaj prelaz se dešava veoma brzo: za oko 10-12 sekundi. Nakon toga, proteinski dio pigmenta također prolazi kroz niz transformacijskih promjena, uz formiranje niza međuprodukata. Jedna od konformacija proteinskog dijela, metarodopsin II, najvažnija je za transdukciju signala (o tome ćemo raspravljati kasnije u ovom poglavlju). Slika 2 prikazuje redoslijed događaja tokom dekolorizacije i regeneracije aktivnog rodopsina. Metahodopsin II se formira nakon 1 ms. Regeneracija pigmenta nakon njegovog raspada se odvija polako, u roku od nekoliko minuta; ovo zahtijeva transport retine od fotoreceptora do pigmentnog epitela.
Struktura rodopsina
Na molekularnom nivou, opsin protein se sastoji od 348 aminokiselinskih ostataka, formirajući 7 hidrofobnih zona, od kojih se svaka sastoji od 20-25 aminokiselina, koje čine 7 transmembranskih spirala. N-terminus molekula nalazi se u ekstracelularnom prostoru (tj. unutar diska štapa), a C-terminus se nalazi u citoplazmi.
Fig.1. Struktura rodopsina kralježnjaka ugrađenog u membranu fotoreceptora. Heliks je donekle odmotan kako bi pokazao lokaciju retine (označeno crnom bojom). C - C-terminus, N - N-terminus.
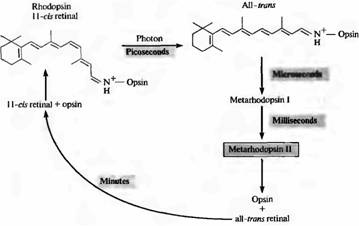
Fig.2. Eflorescencija rodopsina na svjetlu. U mraku, 11-cis-retinal je čvrsto vezan za protein opsin. Hvatanje fotona dovodi do izomerizacije svih cis retinala u retinalnom tronu. U ovom slučaju, opsin all-tron-retinal kompleks brzo se pretvara u metarodopsin II, koji se disocira u opsin i all-tron retinal. Regeneracija rodopsina zavisi od interakcije fotoreceptora i pigmentnih ćelija. Metarhodopsin II aktivira i održava sistem drugog glasnika.
Retinal je povezan sa opsinom preko ostatka lizina koji se nalazi u sedmom transmembranskom segmentu. Opsin pripada porodici proteina sa 7 transmembranskih domena, što uključuje i metabotropne medijatorne receptore, kao što su adrenergički i muskarinski receptori. Poput rodopsina, ovi receptori signaliziraju sekundarnim glasnicima putem aktivacije G-proteina. Rodopsin je izuzetno stabilan u mraku. Bayor je izračunao da spontana termička izomerizacija molekula rodopsina traje oko 3000 godina, ili 10 23 više nego za fotoizomerizaciju.
Konusi i vid boja
Neverovatna istraživanja i eksperimenti koje su sproveli Young i Helmholtz u 19. veku skrenula su pažnju na samu važno pitanje o vidu boja, a sami naučnici dali su jasno i tačno objašnjenje ovog fenomena. Njihov zaključak o postojanju tri razne vrste fotoreceptori u boji su izdržali test vremena i naknadno su potvrđeni na molekularnom nivou. Opet, možemo citirati Helmholtza, koji je uporedio percepciju svjetla i zvuka, boje i zvučnog tona. Može se pozavidjeti na jasnoći, snazi i ljepoti njegove misli, posebno u poređenju sa zbunjujućim vitalističkim konceptima koji su bili rašireni u 19. stoljeću:
Sve razlike u tonovima boja zavise od kombinacije u različitim omjerima tri osnovne boje...crvene, zelene i ljubičaste... Kao što percepcija sunčeve svjetlosti i njene topline ovisi...od toga da li sunčeve zrake pogađaju nervi, koji dolaze iz receptora vida ili iz receptora toplinske osjetljivosti. Kao što je Young sugerirao u svojoj hipotezi, razlika u percepciji različitih boja ovisi jednostavno o tome koji od 3 tipa fotoreceptora je više aktiviran ovom svjetlošću. Kada su sve tri vrste jednako uzbuđena, postaje bela...
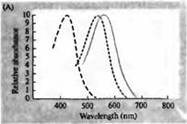
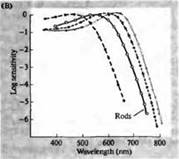
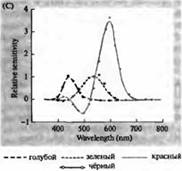
Rice. 3. Spektri osjetljivosti ljudskih fotoreceptora i razni vizuelni pigmenti. (A) Krivulje osjetljivosti tri vizualna pigmenta u boji koje pokazuju vrhunce apsorpcije na valnim dužinama koje odgovaraju cijan, zelenoj i crvenoj. (B) Spektri osjetljivosti čunjeva na plavu, zelenu i crvenu, i štapića (prikazano crnom) kod makaka. Odgovori su zabilježeni korištenjem usisnih elektroda, usrednjeni i normalizirani. Krive spektra štapića dobijene su u proučavanju vidnih pigmenata kod ljudi. (C) Poređenje spektra konusa majmuna i čovjeka korištenjem testa osjetljivosti boja. Kontinuirana krivulja pokazuje eksperiment za određivanje osjetljivosti na boju kod ljudi, kada je predstavljena svjetlošću različitih valnih dužina. Isprekidana linija prikazuje rezultate predviđene na osnovu registracije struja u pojedinačnim čunjićima, nakon korekcije za apsorpciju svjetlosti u sočivu i pigmenta na putu do vanjskog segmenta. Slaganje između rezultata oba eksperimenta je iznenađujuće visoko.
Ako projektujemo dva snopa svjetlosti različitih boja u isto vrijeme na bijeli ekran... vidimo samo jednu boju, manje-više različitu od obje ove boje. Možemo bolje razumjeti izvanrednu činjenicu da smo u mogućnosti da percipiramo sve nijanse u sastavu vanjskog svjetla mješavinom tri osnovne boje, ako uporedimo oko sa suhim... U slučaju zvuka... mi čuje duže talase kao niske tonove, a kratke talase - kao visoke i prodorne, osim toga, uho je u stanju da uhvati više zvučnih talasa u isto vreme, tj. mnoge beleške. Međutim, one β se u ovom slučaju ne spajaju u jedan složen akord, kao što se različite boje ... spajaju u jednu složenu boju. Oko ne može uočiti razliku ako zamijenimo narandžasta boja do crvene ili žute; ali ako čujemo da note do i mi zvuče u isto vrijeme, takav zvuk nam se ne čini kao nota re. Ako uho percipira muzičke tonove kao što oko percipira boje, svaki akord bi mogao biti predstavljen kombinacijom tri konstantne note, jedne vrlo niske, jedne vrlo visoke i jedne srednje, proizvodeći sve moguće muzičke efekte samo promjenom relativne glasnoće ovih tonova. tri note... Međutim, u stanju smo da vidimo glatki prelaz boja iz jedne u drugu kroz beskonačan broj nijansi i gradacija... Način na koji percipiramo svaku od boja... zavisi uglavnom od strukture našeg nervni sistem. Mora se priznati da trenutno ni kod ljudi ni kod tetrapoda nije opisana anatomska osnova koja bi potvrdila teoriju percepcije boja.
Ova precizna i dalekovidna predviđanja potvrđena su nizom različitih zapažanja. Koristeći spektrofotometriju, Wald, Brouck, McNicol i Dartnell et al su pokazali prisustvo tri tipa čunjeva sa različitim pigmentima u ljudskoj mrežnjači. Baylor i kolege su također uspjeli da skrenu struje sa čunjeva majmuna i ljudi. Utvrđeno je da tri populacije konusa imaju različite, ali preklapajuće opsege osjetljivosti na plavi, zeleni i crveni dio spektra. Optimalne talasne dužine za pobuđivanje električnih signala tačno su se poklapale sa vrhovima apsorpcije svetlosti vizuelnim pigmentima, utvrđenim spektrofotometrijom i psihofizičkim eksperimentima za merenje osetljivosti oka na spektar boja. Na kraju, Natais je klonirala i sekvencirala gene koji kodiraju opsin pigment u tri vrste čunjeva koji su osjetljivi na crvenu, zelenu i plavu.
Kako su onda molekuli različitih vizuelnih pigmenata sposobne preferencijalno uhvatiti svjetlost određene valne dužine? Ispostavilo se da je rodopsin vizuelni pigment štapića i da sva tri vizuelna pigmenta čunjeva sadrže isti hromofor, 11-cis-retinal. Međutim, sekvence aminokiselina proteinskog dijela pigmenta razlikuju se jedna od druge. Razlike u samo nekoliko aminokiselina objašnjavaju njihovu različitu osjetljivost na spektar.
daltonizam
Iako jedan tip fotoreceptora nije sposoban da percipira boju sam po sebi, tri tipa čunjića, kao što je prikazano na sl. 4, već sposoban.
U principu, dvije vrste čunjeva s različitim pigmentima bile bi dovoljne za prepoznavanje boja, ali bi se u ovom slučaju brojne kombinacije valnih dužina percipirale podjednako. Slična situacija se događa kada osoba pati od sljepoće za boje. Takvi ljudi, kako je pokazao Nathans, imaju genetske defekte koji dovode do odsustva jednog od pigmenata. Sa stanovišta sadašnjeg stanja nauke, ne možemo a da ne budemo začuđeni koliko lijepo molekularni mehanizmi potvrđuju briljantno i iznenađujuće tačno razmišljanje Younga i Helmholtza.
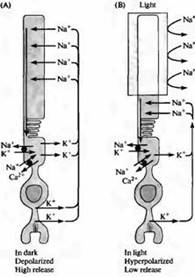
Rice. 4. "Tamna" struja u štapu. (A) U mraku, joni natrijuma prolaze kroz kationske kanale u vanjskom segmentu štapića, uzrokujući depolarizaciju; joni kalcijuma također mogu proći kroz ove katjonske kanale. Strujna petlja prolazi kroz prevlaku štapa zbog činjenice da struja kalija teče prema van u unutrašnjem segmentu membrane. (B) Kada je vanjski segment osvijetljen, kanali se zatvaraju zbog smanjenja intracelularne koncentracije cGMP-a i štap se hiperpolarizira. Hiperpolarizacija dovodi do smanjenja oslobađanja neurotransmitera. Koncentracije natrijuma, kalija i kalcija održavaju se unutar štapa posebnim pumpama i ionskim izmjenjivačima koji se nalaze u području unutrašnjeg segmenta (crni kružići). Transporteri kalcijuma se takođe nalaze u spoljašnjem segmentu.
Njihovu ideju da se glavni atributi daltonizma i sljepoće za boje treba pronaći u samim fotoreceptorima potvrđena je direktnim fiziološkim mjerenjima, kao i proučavanjem razlika u strukturi pigmenata na genetskom i proteinskom nivou, potencijal je reda veličine. od -40 mV, što je daleko od ravnotežnog potencijala kalijuma E K, koji iznosi - 80 mV.

Rice. 5. Uloga cGMP u otvaranju natrijumskih kanala u vanjskom segmentu štapića. Aktivnost pojedinačnih kanala je snimljena pomoću patch stezaljke iznutra prema van, koji su bili u rastvoru sa različitim koncentracijama cGMP. Otvaranje kanala uzrokuje odstupanje struje naviše. Frekvencija otvaranja kanala bila je vrlo niska u kontroli, dodavanje cGMP je dovelo do povećanja frekvencije otvaranja jednog kanala, što je direktno ovisilo o koncentraciji cGMP.
Dolaznu struju u mraku prenose prvenstveno joni natrijuma prateći elektrohemijski gradijent kroz katjonske kanale vanjskog segmenta. Hiperpolarizacija fotoreceptora pod dejstvom svetlosti nastaje zbog zatvaranja ovih kanala, što dovodi do pomeranja membranskog potencijala prema E K.
Svojstva fotoreceptorskih kanala
Kationski kanali spoljašnjeg segmenta u fiziološkim uslovima imaju odnos u provodljivosti jona kalcijuma/natrijuma/kalijuma 12,5:1,0:0,7, a provodljivost jednog kanala je oko 0,1 pS 21). Budući da je koncentracija natrijuma mnogo veća od koncentracije kalcija, oko 85% dolazne struje čine joni natrijuma. Pokretačka snaga jona kalijuma usmjerena je izvan ćelije. Kada ioni kalcijuma prolaze kroz kanal, ovi ioni su čvrsto vezani za zid pora i tako ometaju provodljivost drugih jona. Upravo zbog toga uklanjanje kalcija iz ekstracelularnog medija dovodi do lakšeg prolaska jona kalija i natrijuma kroz kanale, čija se provodljivost povećava na 25 pS.
Fesenko, Yau, Baylor, Strayer i kolege su pokazali da ciklički HMP igra ulogu unutrašnjeg nosača signala od diska do površine membrane. Kao što je prikazano na slici 4, visoka koncentracija cGMP u citoplazmi drži kationske kanale otvorenim. Sa smanjenjem koncentracije cGMP sa unutrašnje površine membrane, otvaranje kationskih kanala postaje rijedak događaj. Dakle, membranski potencijal fotoreceptora je odraz koncentracije cGMP u citoplazmi: što je veća koncentracija cGMP, veća je depolarizacija ćelije. Koncentracija cGMP-a, pak, ovisi o intenzitetu upadne svjetlosti. Povećanje intenziteta svjetlosti dovodi do smanjenja koncentracije cGMP i smanjuje udio otvorenih kanala. U nedostatku cGMP-a, gotovo svi kanali su zatvoreni, a otpornost membrane vanjskog segmenta približava se otporu lipidnog dvosloja.
Molekularna struktura cGMP-gated kanala
cDNA je izolirana za jonske kanale vanjskog segmenta štapića i određene su aminokiselinske sekvence podjedinica ovih kanala u mrežnici čovjeka, goveda, miša i piletine. Pronađena je značajna sličnost DNK za ove i druge cGMP-zavisne jonske kanale - poput onih koji se nalaze u olfaktornim
Transdukcija signala u fotoreceptorima
Kako fotoizomerizacija rodopsina dovodi do promjene membranskog potencijala? Tokom godina se shvatilo da je potrebna neka vrsta unutrašnjeg posrednika za pobuđivanje električnih signala u štapovima i čunjevima. Jedan od razloga za sumnju da se informacija o apsorpciji fotona u području vanjskog segmenta štapića prenosi putem nosača bila je činjenica da se sam pigment rodopsin nalazi unutar diska, a signal se širi kroz citoplazmu do spoljnu membranu. Drugi razlog je bio značajan porast odziva. Baylor i kolege, proučavajući fotoreceptore u kornjačama, pokazali su da se smanjenje membranske provodljivosti i zabilježenih električnih signala događa već kada se apsorbira samo jedan foton i aktivira jedan od 10 8 pigmentnih molekula.
Slijed događaja u kojima aktivirana molekula fotopigmenta mijenja membranski potencijal razjašnjen je proučavanjem vanjskih segmenata štapića i čunjeva korištenjem stezanja potencijala s dvije elektrode i metodama molekularne biologije. Šema prijenosa signala od apsorpcije fotona svjetlosti do električnog signala prikazana je na sl. 3.
U mraku, u vanjskim segmentima štapića i čunjeva teče stalna dolazeća "tamna" struja. Rezultat je njihov membranski sistem. Područja membrane ovih kanala pokazuju strukturne sličnosti sa drugim kationsko-selektivnim kanalima, posebno u S4 regiji i u regiji koja formira jonske pore. Fotoreceptorski jonski kanali su tetrameri sastavljeni od najmanje 2 različite proteinske podjedinice s molekulskom težinom od 63 odnosno 240 kDa.
Intracelularna mjesta vezanja nukleotida nalaze se blizu karboksilnog kraja podjedinica. Ekspresija ovih podjedinica u oocitima dovodi do stvaranja katjonskih kanala koji imaju svojstva slična kanalima koji se nalaze u vanjskim segmentima štapića: aktiviraju se cGMP i imaju očekivane omjere provodljivosti i permeabilnosti.

Rice. 6. Mehanizam aktivacije G-proteina nakon aktivacije molekula fotopigmenta. G-protein transducin vezuje GTP u prisustvu metarodopsina II, što dovodi do aktivacije fosfodiesteraze, koja zauzvrat hidrolizira cGMP. Kada koncentracija cGMP padne, natrijumski kanali se zatvaraju.
Metabolička kaskada cikličkog GMP-a
Lanac događaja koji dovode do smanjenja koncentracije cGMP i naknadnog zatvaranja jonskih kanala prikazan je na slici 5. Smanjenje intracelularne koncentracije cGMP uzrokovano je svjetlom, što dovodi do stvaranja metarodopsina II, međuproizvoda u razgradnji fotopigmenta. Metarhodopsin II zauzvrat djeluje na G-protein transducin, koji se sastoji od 3 polipeptidna lanca
Interakcija metarodopsina II i transducina dovodi do zamjene molekula GDP-a vezanog za G-protein GTP-om. Ovo aktivira podjedinicu G-proteina, koja se odvaja od podjedinica i, zauzvrat, aktivira fosfodiesterazu koja se nalazi u membranskoj regiji: enzim koji hidrolizira cGMP. Koncentracija cGMP opada, ima manje otvorenih ionskih kanala, štap postaje hiperpolariziran. Kaskada je prekinuta fosforilacijom C-terminusa aktivnog metarodopsina II. Ključna uloga cGMP-a u regulaciji stanja katjonskih kanala potvrđena je biohemijskim eksperimentima. Osvetljenje fotoreceptora može izazvati pad nivoa cGMP u ćeliji za 20%.
Receptori kralježnjaka koji se depolariziraju pri izlaganju svjetlosti
Zanimljiv izuzetak od gore navedenog mehanizma fotorecepcije su neki receptori kičmenjaka. Gušteri imaju treće oko koje se nalazi na vrhu glave. Sadrži male "čušture" sposobne da percipiraju sliku sličnu onoj koju percipiraju glavne (bočne) oči. Ovi fotoreceptori su, međutim, izvanredni po tome što se depolariziraju nakon osvjetljenja. Nukleotidski vođeni kanali ovdje imaju sličnu strukturu i funkciju kao fotoreceptori drugih kralježnjaka, s jednim izuzetkom: aktivacija fotoreceptora i G-proteina dovodi do povećanja koncentracije cGMP. Kao rezultat, kanali vanjskog segmenta se otvaraju, a kationi se usmjeravaju u ćeliju, formirajući "svjetlu" struju. To je zbog inhibicije aktivnosti fosfodiesteraze u mraku. Kao rezultat, u oku guštera dolazi do sljedećeg slijeda događaja: svjetlost - [cGMP] - otvaranje katjonskih kanala vanjskog segmenta - depolarizacija.
Pojačanje signala u cGMP kaskadi
Dvostepena cGMP kaskada obezbeđuje značajno pojačanje originalnog signala, što objašnjava izuzetnu osetljivost štapova na svetlost. Prvo, jedan molekul aktivnog metarodopsina II katalizira dodavanje mnogih GTP molekula umjesto GDP-a i tako oslobađa stotine G-proteinskih podjedinica. Drugo, svaka podjedinica aktivira jedan molekul fosfodiesteraze u disku, koji je u stanju da cijepa ogroman broj cGMP molekula koji se nalaze u citoplazmi i tako dovede do zatvaranja velikog broja kanala.
Signali kao odgovor na pojedinačne kvante svjetlosti
Podaci da pojedinačni kvanti svjetlosti mogu uzrokovati opaženi osjećaj svjetlosti veliki broj pitanja. Koliki je ovaj pojedinačni odgovor? Kako se ovaj signal izdvaja od nivoa buke? I kako se takve informacije pouzdano prenose od mrežnice do viših vidnih centara? Kako bi izmjerili signale kao odgovor na pojedinačne kvante svjetlosti, Baylor i kolege snimili su struje iz pojedinačnih štapića u mrežnjačima žaba, majmuna i ljudi. Ovi eksperimenti su jedinstven primjer eksperimenta koji pokazuje kako se tako složen proces kao što je percepcija slabih bljeskova svjetlosti može povezati s promjenama koje se dešavaju na nivou pojedinačnih molekula.
Postupak izolacije fragmenta retine iz životinjskog ili leševnog materijala treba provesti u mraku za ove eksperimente. Da bi se izmjerila struja, vanjski segment štapića se usisava u tanku pipetu. Kao što se i očekivalo, ovi eksperimenti pokazuju da u mraku struja stalno teče unutar vanjskog segmenta. Bljeskovi svjetlosti zatvaraju kanale u vanjskom segmentu, uzrokujući smanjenje "tamne" struje. Amplituda struje je mala i proporcionalna broju apsorbovanih fotona. Ponekad blic uzrokuje jednostruki odgovor, ponekad dvostruki odgovor, a ponekad nikakav odgovor.
Kod majmunskih štapova, smanjenje struje kao odgovor na apsorpciju jednog fotona iznosi oko 0,5 pA. To odgovara zatvaranju oko 300 kanala, tj. od 3 do 5% svih kanala se otvara u mraku. To se postiže značajnim povećanjem signala u metaboličkom sustavu cGMP kaskada. Štaviše, zbog ekstremne stabilnosti vizuelnih pigmenata spomenutih ranije, slučajna izomerizacija i lažno zatvaranje kanala su vrlo rijetki događaji. Ovo uzrokuje da se efekti pojedinačnih svjetlosnih kvanta ističu na pozadini vrlo niske istosmjerne buke. Pokazalo se da električna sprega između fotoreceptora pruža dodatni efekat izglađivanja koji smanjuje pozadinsku buku i poboljšava omjer signala i šuma odgovora štapa na pojedinačne kvante.
Književnost
1. Finn, J. T., Grunwald, M. E, i Yau, K-W. 1996. Ciklični nukleotidni ionski kanali: proširena porodica sa različitim funkcijama. Annu. Rev. Physiol.58: 395-426.
2. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y, Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S., i Okada, M. 1998. Glutamatni receptori : Funkcija mozga i transdukcija signala.
VIZUELNI PIGMENTI(lat. pigmentum paint) - fotosenzitivni pigmenti fotoreceptora retine. Opažajući energiju svetlosnog impulsa, 3. predmeti prolaze kroz složen fotohemijski ciklus. transformacije, kao rezultat kojih odvojeni vidni receptor retine, koji sadrži 3. p. (konus ili štapić), prelazi u pobuđeno stanje i prenosi primljenu informaciju duž optičkog živca do c. n. With. Budući da su glavni strukturni i funkcionalni dio fotoreceptorske membrane vizualnih stanica retine oka, 3. predmeti igraju ključnu ulogu u mehanizmima vida (vidi).
Nomenklatura i struktura vizuelnih pigmenata. Svi proučavani 3. p. kičmenjaka i beskičmenjaka su kompleksi membranskog proteina opsina netopivog u vodi i hromofora (retinala) koji je povezan s njim. Retinal, ili aldehid vitamina A, može postojati u dva oblika, retinal1 i retinal2.
Po prirodi hromofora 3. stavka je podijeljena u dvije klase - rodopsine (vidi), koji sadrže retinal1, i porfiropsine koji sadrže retinal2. Rodopsini se nalaze u retini oka svih kopnenih i morskih životinja, porfiropsini se nalaze u retini očiju slatkovodnih životinja. Neke ribe i vodozemci imaju 3 p. koji sadrže retinal u isto vrijeme! i retina. Postoje pokušaji da se klasifikuje 3. p. na osnovu razlika u opsinima specifičnim za štapiće ili čunjiće retine. Na primjer, rodopsin je kompleks retinal1 sa šipkastim opsinom, jodopsin je kompleks retinal1 sa opsinom konusa, porfiropsin je retinal2 sa šipkastim opsinom, kompleks retinal-konus opsina formira cijanopsin. Međutim, izuzetno je teško klasifikovati 3. stavke na osnovu opsina, jer postoji najmanje pet različitih opsina.
Od svih poznatih 3. predmeta, najpotpunije su istraženi rodopsini izolovani iz očiju bika, žabe i lignje. Njihova molba. težine (mase) reda 30-40 hiljada, svaki molekul sadrži cca. 400 aminokiselina i jedan hromofor. Osim toga, oligosaharidni lanac je uključen u molekulu 3. p.: 3 glukozaminska radikala, 2 manoza, 1 galaktoza. Lipidi (ch. arr. fosfolipidi) formiraju jak kompleks sa molekulom 3. p. Zadržavajući svoja osnovna spektralna svojstva (vidi spektralnu analizu), 3. artikli bez lipida gube niz funkcionalno važnih, na primjer, sposobnost oporavka.
Čista retina ima žuta, maksimum njegovog apsorpcionog spektra leži u području od 370 nm. Opsin je bezbojan, maksimum apsorpcije je u ultraljubičastom području (cca. 280 nm). Boja molekule rodopsina je crvenkasto-ružičasta, maksimalni apsorpcioni spektar je cca. 500 nm. Razlog tako snažnog spektralnog pomaka tokom formiranja kompleksa (od 370 do 500 nm - tzv. batohromski pomak) još nije dobio jednoznačno objašnjenje.
Maksimumi apsorpcionih spektra rodopsina i porfiropsina obuhvataju prilično široku oblast vidljivog spektra - od 433 do 562 nm za rodopsine i od 510 do 543 nm za porfiropsin. Ako porfiropsin uključuje i 3.p. čunjeve punoglavca žabe, šarana i slatkovodne kornjače, odnosno cijanopsin sa maksimalnim spektrom apsorpcije na 620 nm, onda je ovo područje još šire. Razvoj metoda mikrospektrofotometrije omogućio je određivanje apsorpcionih spektra mnogih tipova pojedinačnih fotoreceptorskih ćelija kod životinja i ljudi. Prema dobijenim podacima, 3. p. ljudske retine imaju sledeće maksimume apsorpcionog spektra: štapići 498, čepići osetljivi na plavo, zeleno i crveno - 440, 535 i 575 nm, respektivno.
Studij 3. str je započeo na njemačkom jeziku. istraživač H. Muller, koji je 1851. opisao kako ružičasto-ljubičasta mrežnica izvađena iz oka žabe postaje prvo žućkasta na svjetlu, a zatim bjelkasta. F. Boll je 1877. godine također opisao ovu pojavu, zaključivši da se neka vrsta tvari osjetljive na crveno svjetlo nalazi u vidnim ćelijama mrežnjače i da je promjena boje ove supstance povezana sa mehanizmom vida. Velika zasluga u proučavanju 3. p. pripada Kuhneu (W. Kuhne, 1877), Krom je uspio izolovati 3. p. i detaljno ih proučiti. On je 3. p. koji je izdvojio nazvao vizuelno ljubičastim, utvrdio njegovu proteinsku prirodu, istražio neka od njegovih spektralnih svojstava i fototransformacija, otkrio sposobnost 3. p. da se oporavlja u mraku. Amer je dao veliki doprinos proučavanju 3. str. biohemičar i fiziolog J. Wald.
Fototransformacije vidnih pigmenata. Pod uticajem svetlosti na 3. p. u njima nastaje fotohemijski ciklus. transformacije, koje se zasnivaju na primarnoj fotohemijskoj reakciji cis-trans izomerizacije retine (vidi Izomerizam). U tom slučaju dolazi do prekida veze između hromofora i proteina. Slijed transformacija 3. p. može se predstaviti na sljedeći način: rodopsin (hromofor je u cis obliku) -> prelumirhodopsin -> lumirhodopsin -> metarhodopsin I -> metarhodopsin II -> opsin protein -> hromofor u trans obliku . Pod utjecajem enzima - retinol dehidrogenaze - potonji prelazi u vitamin A, koji dolazi iz vanjskih segmenata štapića i čunjića u ćelije pigmentnog sloja retine. Prilikom potamnjivanja oka dolazi do regeneracije 3. p., za izvođenje reza neophodno je prisustvo cis-izomera vitamina A, koji služi kao polazni proizvod za stvaranje hromofora (vitamin A aldehid). . Uz nedostatak ili odsustvo vitamina A u organizmu, formiranje rodopsina može biti poremećeno i razviti se kao rezultat poremećaja vida u sumrak, tzv. noćno sljepilo (vidi Hemeralopia). U procesu fototransformacije rodopsina u fazi tranzicije lumirhodopsina u metarodopsin I u receptorskoj ćeliji nastaje kao odgovor na sjajni bljesak tzv. potencijal ranog (kratke latencije) receptora. Međutim, nije vizuelni signal, iako može poslužiti kao jedan od testova za proučavanje mehanizma transformacija 3. p. u membrani fotoreceptora. Funkcionalna vrijednost je tzv. kasnog receptorskog potencijala, latentni period to-rogo (5-10 msec) je proporcionalan vremenu formiranja metarodopsina II. Pretpostavlja se da reakcija tranzicije metarodopsina I u metarodopsin II obezbeđuje pojavu vizuelnog signala.
3. Budući da se predmeti kontinuirano mijenjaju na svjetlu, moraju postojati mehanizmi za njihovu stalnu restauraciju. Neki od njih su izuzetno brzi (fotoregeneracija), drugi su dosta brzi (biohemijski, regenerativni, tamni), treći su spori (sinteza 3. p. Tokom stalnog obnavljanja fotoreceptorske membrane u vidnoj ćeliji). Fotoregeneracija ima fiziolnu vrijednost kod beskičmenjaka (na primjer, kod glavonošci- lignje, hobotnica). U mehanizmu biohemije. regeneracija 3. p. kod kičmenjaka, važnu ulogu, očigledno, igra enzim izomeraza (vidi), koji obezbeđuje izomerizaciju trans-retinala (ili trans-vitamina A) ponovo u cis-izomerni oblik. Međutim, ne postoje definitivni dokazi za postojanje takvog enzima. Ista reakcija formiranja molekula od 3. p. u prisustvu 11-cis-izomera retine i opsina u sistemu se odvija lako, bez utroška energije. Pronađena je sposobnost izbijeljenog rodopsina na reakciju fosforilacije (vidi); pretpostavlja se da je ova reakcija jedna od karika u mehanizmu svjetlosne adaptacije vidne ćelije.
Bibliografija: Ackerman Yu. Biophysics, trans. sa engleskog, M., 1964; Willie K. i Det e V. Biology, trans. iz engleskog, M., 1974, bibliografija; Konev S. V. i Volotovsky I. D. Uvod u molekularnu fotobiologiju, str. 61, Minsk, 1971; Ostrovsky M. A. i Fedorovich I. B. Fotoinducirane promjene na membrani fotoreceptora, u knjizi: Struktura i funkcije biola, membrane, ur. A. S. Troshina i drugi, str. 224, M., 1975, bibliogr.; Fiziologija senzornih sistema, ur. G. V. Gershuni, dio 1, str. 88, L., 1971; Biohemija i fiziologija vidnih pigmenata, ur. od H. Langera, B. a. o., 1973; Priručnik za senzornu fiziologiju, ur. od H. A. R. Jung a. o., v. 7, pt 1-2, B., 1972.
M. A. Ostrovsky.