pigmenți vizuali. fotorecepție. Care dintre cochiliile globului ocular este transparentă în față. Corpul geniculat lateral este o structură
PIGMENTI VIZUALI(lat. pigmentum paint) - pigmenți fotosensibili ai fotoreceptorilor retinei. Percepând energia unui impuls de lumină, 3. elementele suferă un ciclu fotochimic complex. transformări, în urma cărora un receptor vizual separat al retinei, care conține 3. p. (un con sau o tijă), intră într-o stare excitată și transmite informațiile primite de-a lungul nervului optic către c. n. Cu. Fiind principala parte structurală și funcțională a membranei fotoreceptoare a celulelor vizuale ale retinei, 3. p. Rol cheieîn mecanismele vederii (vezi).
Nomenclatură și structură pigmenți vizuali. Toate 3. p. studiate ale vertebratelor și nevertebratelor sunt complexe ale proteinei membranare insolubile în apă opsina și cromoforul (retinianul) asociat acesteia. Aldehida retiniană sau vitamina A poate exista în două forme, retinină1 și retinină2.
Prin natura unui cromofor 3. itemul este împărțit în două clase - rodopsine (vezi), care conțin retina1 și porfiropsinele care conțin retina2. Rodopsinele se găsesc în retina ochiului tuturor animalelor terestre și marine, porfiropsinele se găsesc în retina ochilor animalelor de apă dulce. Unii pești și amfibieni au 3 p. care conțin retina în același timp! și retiniană. Există încercări de clasificare a 3. p. pe baza diferențelor de opsine specifice bastonașelor sau conurilor retinei. De exemplu, rodopsina este un complex de retina1 cu opsina de tijă, iodopsina este un complex de retina1 cu opsina de con, porfiropsina este retina2 cu opsina de tijă, complexul de opsina de retinină-con formează cianopsina. Cu toate acestea, este extrem de dificil să 3. clasificăm articolele pe baza opsinelor, deoarece există cel puțin cinci opsins diferite.
Dintre toate 3. elementele cunoscute, rodopsinele izolate din ochii unui taur, broaște și calmar au fost investigate cel mai complet. Pledoaria lor. greutate (masă) de ordinul a 30-40 mii, fiecare moleculă conține cca. 400 de aminoacizi și un cromofor. În plus, lanțul oligozaharidic este inclus în molecula de 3. p.: 3 radicali de glucozamină, 2 manoză, 1 galactoză. Lipidele (ch. arr. fosfolipide) formează un complex puternic cu molecula 3. p. În timp ce își păstrează proprietățile spectrale de bază (vezi Analiza spectrală), 3. elementele fără lipide își pierd o serie de elemente importante din punct de vedere funcțional, de exemplu, capacitatea de a se recupera.
Retina pură are o culoare galbenă, maximul spectrului său de absorbție se află în regiunea de 370 nm. Opsina este incoloră, maximul de absorbție este în regiunea ultravioletă (aprox. 280 nm). Culoarea moleculei de rodopsina este roz-roșcat, spectrul maxim de absorbție este de cca. 500 nm. Motivul unei schimbări spectrale atât de puternice în timpul formării complexului (de la 370 la 500 nm - așa-numita schimbare batocromică) nu a primit încă o explicație clară.
Maximele spectrelor de absorbție ale rodopsinei și porfiropsinei captează o regiune destul de largă a spectrului vizibil - de la 433 la 562 nm pentru rodopsine și de la 510 la 543 nm pentru porfiropsine. Dacă porfiropsinele includ și conuri 3p ale mormolocului unei broaște, crap și broaște țestoasă de apă dulce, adică cianopsină cu un spectru maxim de absorbție la 620 nm, atunci această regiune este și mai largă. Dezvoltarea metodelor de microspectrofotometrie a făcut posibilă determinarea spectrelor de absorbție a multor tipuri de celule fotoreceptoare unice la animale și la oameni. Conform datelor obținute, 3. p. ale retinei umane au următoarele maxime de spectru de absorbție: tije 498, conuri sensibile la albastru, verde și roșu - 440, 535 și, respectiv, 575 nm.
Studiul 3. p. a început în limba germană. cercetătorul H. Muller, care în 1851 a descris cum retina roz-violet extrasă din ochiul unei broaște devine mai întâi gălbuie la lumină, apoi albicioasă. În 1877, F. Boll a descris și el acest fenomen, ajungând la concluzia că un fel de substanță roșie sensibilă la lumină este localizată în celulele vizuale ale retinei și că decolorarea acestei substanțe este asociată cu mecanismul vederii. Un mare merit în studiul 3. p. îi aparține lui Kuhne (W. Kuhne, 1877), Krom a reușit să izoleze 3. p. și să le studieze în detaliu. El a numit 3. p. extras de el vizual violet, i-a stabilit natura proteică, a investigat unele dintre proprietățile spectrale și fototransformările sale, a descoperit capacitatea lui 3. p. de a se recupera în întuneric. Amer. a adus o mare contribuţie la studiul 3. p. biochimistul și fiziologul J. Wald.
Fototransformări ale pigmenților vizuali. Sub influența luminii asupra 3. p., în ele are loc un ciclu fotochimic. transformări, care se bazează pe reacția fotochimică primară a izomerizării cis-trans a retinei (vezi Izomerism). În acest caz, legătura dintre cromofor și proteină este întreruptă. Secvența transformărilor lui 3. p. poate fi reprezentată astfel: rodopsina (cromoforul este în formă cis) -> prelumirhodopsin -> lumirhodopsin -> metarhodopsin I -> metarhodopsin II -> opsin protein -> cromofor în formă trans . Sub influența enzimei - retinol dehidrogenaza - aceasta din urmă trece în vitamina A, care vine din segmentele exterioare ale tijelor și conurilor în celulele stratului pigmentar al retinei. La întunecarea ochiului, are loc regenerarea 3. p., pentru implementarea unei tăieturi, este necesară prezența unui izomer cis al vitaminei A, care servește ca produs de pornire pentru formarea unui cromofor (aldehidă de vitamina A). . Cu lipsa sau absența vitaminei A în organism, formarea rodopsinei poate fi perturbată și se poate dezvolta ca urmare a unei tulburări a vederii crepusculare, așa-numita. orbire nocturnă (vezi Hemeralopia). În procesul de fototransformare a rodopsinei în stadiul de tranziție a lumirhodopsinei la metarodopsină I în celula receptoră are loc ca răspuns la o strălucire strălucitoare a așa-numitului. potențial de receptor precoce (cu latență scurtă). În același timp, nu este un semnal vizual, deși poate servi drept unul dintre testele pentru studiul mecanismului de transformări ale 3. p. în membrana fotoreceptorului. Valoarea funcțională este așa-numita. potențial tardiv al receptorului, perioada latentă to-rogo (5-10 msec) este proporțională cu timpul de formare a metarhodopsinei II. Se presupune că reacția de tranziție a metarodopsinei I la metarodopsină II oferă aspectul semnal vizual.
Deoarece 3. articolele sunt continuu decolorate la lumină, trebuie să existe mecanisme pentru restaurarea lor constantă. Unele dintre ele sunt extrem de rapide (fotoregenerare), altele sunt destul de rapide (biochimice, regenerare, întunecate), altele sunt lente (sinteza de 3. p. În timpul reînnoirii constante a membranei fotoreceptoare din celula vizuală). Fotoregenerarea are fiziol, valoare la animalele nevertebrate (de exemplu, la cefalopode- calmar, caracatiță). În mecanismul biochimiei. regenerarea 3. p. la vertebrate, un rol important, aparent, îl joacă enzima izomeraza (vezi), care asigură izomerizarea trans-retinianului (sau trans-vitamina A) din nou în forma de izomer cis. Cu toate acestea, nu există dovezi definitive pentru existența unei astfel de enzime. Aceeași reacție de formare a unei molecule de 3. p. în prezența izomerului 11-cis al retinei și opsinei în sistem are loc cu ușurință, fără cheltuială de energie. Se constată capacitatea rodopsinei albite la reacția de fosforilare (vezi); se presupune că această reacție este una dintre verigile din mecanismul de adaptare la lumină a celulei vizuale.
Bibliografie: Ackerman Yu. Biofizică, trad. din engleză, M., 1964; Willie K. și Det e V. Biologie, trad. din engleză, M., 1974, bibliografie; Konev S. V. și Volotovsky I. D. Introducere în fotobiologia moleculară, p. 61, Minsk, 1971; Ostrovsky M. A. și Fedorovich I. B. Modificări fotoinduse în membrana fotoreceptorului, în cartea: Structura și funcțiile biol, membrane, ed. A. S. Troshina și alții, p. 224, M., 1975, bibliogr.; Fiziologia sistemelor senzoriale, ed. G. V. Gershuni, partea 1, p. 88, L., 1971; Biochimia și fiziologia pigmenților vizuali, ed. de H. Langer, B. a. o., 1973; Manual de fiziologie senzorială, ed. de H. A. R. Jung a. o., v. 7, pct. 1-2, B., 1972.
M. A. Ostrovsky.
pigment vizual unitate structurală și funcțională a membranei sensibile la lumină a fotoreceptorilor (vezi Fotoreceptorii) retinei - tije și conuri. În Z. p., se realizează prima etapă a percepției vizuale - absorbția cuantelor de lumină vizibilă. Molecula Z. (masa molara de aproximativ 40.000) este formata dintr-un cromofor care absoarbe lumina si opsina, un complex de proteine si fosfolipide. Cromoforul tuturor Z. p. este aldehida vitaminei A 1 sau A 2 - retiniană sau 3-dehidroretinală. Două tipuri de opsină (tijă și con) și două tipuri de retinină, atunci când sunt combinate în perechi, formează 4 tipuri de z.p. nm), iodopsină (562 nm), porfiropsină (522 nm) și cianopsină (620 nm). Legătura fotochimică primară în mecanismul vederii (Vezi Viziunea) constă în fotoizomerizarea retinei, care, sub acțiunea luminii, își schimbă configurația curbă într-una plată. Această reacție este urmată de un lanț de procese întunecate care duc la apariția unui semnal receptor vizual, care este apoi transmis sinaptic la următoarele elemente nervoase ale retinei - celulele bipolare și orizontale. Lit.: Fiziologia sistemelor senzoriale, partea 1, L., 1971, p. 88-125 (Manual de fiziologie); Wald G., Baza moleculară a excitației vizuale, „Natura”, 1968, v. 219. M. A. Ostrovsky.
Marea Enciclopedie Sovietică. - M.: Enciclopedia Sovietică. 1969-1978 .
Vedeți ce este „pigmentul vizual” în alte dicționare:
Funcțional structural. unitate sensibilă la lumină. membranele fotoreceptoare de baston și con din retină. Molecula 3. p. constă dintr-un cromofor care absoarbe lumina și o opsină dintr-un complex de proteine și fosfolipide. Cromoforul este reprezentat de vitamina A1 aldehida ...... Dicționar enciclopedic biologic
Rodopsina (violet vizual) este principalul pigment vizual din tijele retinei umane și animale. Se referă la proteine complexe cromoproteine. Modificările proteinelor caracteristice diferitelor specii biologice pot varia semnificativ... Wikipedia
PIGMENT(E) VIZUAL(E)- Vezi fotopigment... Dicţionarîn psihologie
Pigmentul retinian conținut în interiorul tijelor, care include vitamina A (retiniană) și proteine. Prezența rodopsinei în retină este necesară pentru vederea normală în lumină slabă. Sub influența luminii ...... termeni medicali
RHODOPSINĂ (RHODOPSINĂ), VISUAL MOV- (vizual violet) pigment retinian conținut în interiorul bastoanelor, care include vitamina A retiniană (retiniană) și proteine. Prezența rodopsinei în retină este necesară pentru vederea normală în lumină slabă. Sub… … Dicţionar explicativ de medicină
- (violet vizual), fotosensibil. proteine complexe, pigmentul vizual al celulelor bastonașe din retina vertebratelor și oamenilor. Absorbind o cantitate de lumină (absorbție maximă aprox. 500 nm), R. se dezintegrează și provoacă excitație ... ... Științele naturii. Dicţionar enciclopedic
- (pigment vizual), proteina bastonașă sensibilă la lumină a retinei vertebratelor și celulelor vizuale ale nevertebratelor. R. glicoproteina (mol. m. aprox. 40 mii; lanțul polipeptidic este format din 348 de resturi de aminoacizi), care conține ... ... Enciclopedia chimică
- (din grecescul rhódon rose și ópsis vision) violet vizual, principalul pigment vizual al tijelor retinei vertebratelor (cu excepția unor pești și amfibieni în stadiile incipiente de dezvoltare) și nevertebrate. Conform substanței chimice ...... Marea Enciclopedie Sovietică
- (violet vizual), o proteină complexă sensibilă la lumină, principalul pigment vizual al celulelor bastonașe ale retinei la vertebrate și oameni. Absorbind o cantitate de lumină (absorbția maximă este de aproximativ 500 nm), rodopsina se descompune și provoacă ... ... Dicţionar enciclopedic
Articolul principal: Tije (retină) Rhodopsin (un nume învechit, dar încă folosit vizual violet) este pigmentul vizual principal. Conținut în bastoanele retinei ochiului nevertebratelor marine, peștilor, aproape toate terestre ...... Wikipedia
25-10-2014, 13:04
Quantele de lumină sunt absorbite în receptori de molecule specializate - fotopigmente vizuale. Pigmenții vizuali au fost descoperiți independent de fiziologii germani F. Boll și W. Kuehne în 1877-1879. F. Boll a observat că retina izolată de sticla vizuală a unei broaște la început pare roșie, apoi, estompând în lumină, devine galbenă și, în cele din urmă, complet incoloră.
W. Kuehne a descoperit că într-un ochi al unui animal care funcționează în mod normal, culoarea retinei după iluminarea intensă cu lumină este restabilită dacă animalul este plasat din nou în întuneric. Pe baza acestor observații s-a ajuns la concluzia că în retina ochiului există o substanță fotosensibilă (pigment), a cărei concentrație scade la lumină și se restabilește în întuneric.
pigmenți vizuali- acestea sunt molecule complexe de cromolipoproteine, care la vertebrate și nevertebrate constau din două părți principale: un cromofor (partea colorantă a moleculei care determină culoarea receptorului când este iluminată) și o proteină (opsina). Cromoforul determină maximul și intensitatea absorbției luminii în pigment și este o aldehidă a alcoolilor - vitaminele A1 și A2.
Numele acestor aldehide sunt retina-1 și retina-2. Prezența unei duble legături suplimentare în retina-2 duce la o deplasare a maximului spectrului de absorbție către regiunea cu lungime de undă lungă. Retina se atașează de opsină, o mutație a proteinei care se găsește numai în fotoreceptori. Tija opsina, scotopsina, și conurile, fotopsina, sunt izolate. Scotopsinul constă din 230-270 de resturi de aminoacizi, a căror secvență nu a fost încă determinată.
Se știu puține lucruri despre fotopsină: există doar date despre configurația sa elicoială. Partea actuală care absoarbe lumina a moleculei de pigment, retina, este un tip de pigment carotenoid, cel mai comun în fotosintetic. țesuturi de plante și animale.
Caracteristicile spectrale ale pigmenților sunt determinate de combinații ale uneia dintre retine cu una sau alta varietate de opsine, a căror diversitate determină diversitatea pigmenților vizuali la diferite animale. Astfel, combinarea scotopsinei cu retina-1 duce la formarea unui pigment de tije - rodopsina cu un maxim de absorbtie la? = 500 nm, iar combinarea sa cu retina-2 formeaza un alt pigment de tije - porfiropsina cu un maxim de absorbtie la? =525 nm. Combinatii de retina-1 cu alt fel fotopsinele din retina oamenilor şi maimuţelor formează pigmenţi conici cu maxime de absorbţie la? = 445 nm, ? = 535 nm și? \u003d 570 nm, numit cyanolab, chlorolab și, respectiv, erythrolab.
Proprietățile de absorbție a luminii ale pigmenților sunt determinate tocmai de interacțiunea cromoforului de iopsină: maximele de absorbție ale cromoforului și opsina, luate separat, sunt localizate în regiune? \u003d 278 nm (retina-1) și? = 378 nm (scotopsina), in timp ce compusul lor - rodopsina - are un maxim de absorbtie la? = 500 nm. Maximul de absorbție este unul dintre parametrii caracteristicilor spectrale ale pigmentului, care determină, în general, eficiența absorbției de către pigment a cuantelor de lumină (fotoni) de diferite lungimi de undă (Fig. 3.1.4a).
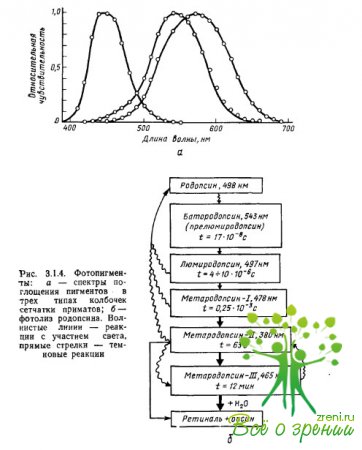
Primele măsurători ale spectrelor de absorbție ale pigmenților în conurile individuale ale retinei maimuțelor și oamenilor au fost efectuate de Brown, Wald și Marx și colab., folosind o metodă de microspectrofotometrie special dezvoltată în acest scop. Preparatul retinian a fost plasat la microscop, ceea ce a făcut posibilă, sub control vizual, focalizarea unui fascicul de lumină de una sau alta lungime de undă pe segmentul exterior al fotoreceptorului, care conține pigmentul.
Apoi, pentru fiecare lungime de undă, s-au făcut măsurători separat ale cantității de lumină care a trecut prin con și nu a fost absorbită de acesta (diferența dintre cantitatea de lumină furnizată conului și cantitatea de lumină transmisă de acesta a caracterizat eficiența absorbția fotonilor cu o lungime de undă dată de către pigment). Măsurătorile au fost repetate după decolorarea pigmentului ca urmare a iluminării intense (măsurători de referință). Ca urmare a scăderii celei de-a doua serii de măsurători din prima, s-a obţinut spectrul diferenţial de absorbţie al pigmentului în funcţie de sensibilitatea la lumină pe lungimea de undă a radiaţiei.
Din fig. 3.1.4a se poate observa că pigmenții au spectre de absorbție de bandă largă, puternic suprapuse. Toți fotopigmenții au un maxim de absorbție suplimentar caracteristic carotenoizilor (așa-numitul nis-peak" sau "?-peak"), egal cu 2/3 din maximul principal. Pentru pigmenți, ale căror maxime principale de absorbție sunt localizate în părțile cu lungime de undă scurtă și medie ale spectrului, vârful cis cade în regiunea ultravioletă și nu afectează analiza radiației din spectrul vizibil. Dacă, în plus, atunci când se construiesc funcțiile sensibilității spectrale a pigmenților, scara lungimii de undă (axa abscisei) este convertită într-o scară de frecvență, atunci spectrele de absorbție ale tuturor pigmenților, inclusiv opsine diferite, dar același rețial (de exemplu, retiniană). -1), va avea aceeași și aceeași formă.
Cu această prezentare de date, propusă pentru prima dată de Dartnall „(1953) pentru pigmenții pe bază de retina-1, spectrele de absorbție ale diferiților fotopigmenți diferă doar în poziția maximului. Și apoi, cunoscând maximul de absorbție al pigmentului, putem restabiliți forma spectrului său de absorbție, mai întâi în coordonatele „frecvența radiației - sensibilitate”, apoi, folosind raportul c = ?f și în coordonatele „lungime de undă - sensibilitate”.
Această metodă de restabilire a spectrului de absorbție al unui pigment necunoscut din date numai la maximul său se numește metoda nomogramei. Metoda nomogramelor este, de asemenea, utilizată pe scară largă în neurofiziologia vederii culorilor atunci când se rezolvă problema numărului și tipului de fotodetectoare care converg către unul sau altul neuron sensibil la culoare: funcția de sensibilitate spectrală a unui neuron este comparată cu una sau alta nomogramă ( sau combinația lor).
Diversitatea fotopigmenților din regnul animal se corelează cu diversitatea habitatelor de mediu. La pești și amfibieni, retina poate conține atât rodopsina, cât și porfiropsină, al căror raport se modifică în timpul vieții individului; la pestii anadromi, inlocuirea rodopsinei cu porfiropsina precede trecerea de la apa sarata la apa dulce si invers. Asocierea compoziției pigmentului cu ecologia habitatului nu este rigidă: modificările compoziției pigmentului pot fi cauzate de schimbări sezoniere și ritmuri circadiene.
Fotoizomerizare.
Reacția unui fotopigment la lumină este determinată de numărul de quante absorbite, indiferent de lungimea de undă și de energia radiației: de exemplu, cele absorbite de rodopsina. 10 cuante de lumină la? \u003d 430 nm provoacă aceleași modificări structurale în ea (Fig. 3.1.46), precum și 10 cuantii de lumină absorbite la? = 500 nm.
Și selectivitatea spectrală (spectrul de absorbție) a pigmentului este determinată doar de probabilitatea diferită de absorbție a cuantelor de lumină din diferite părți ale spectrului vizibil. Astfel, pigmenții vizuali și, în cele din urmă, fotoreceptorii, funcționează ca „contoare de fotoni ai luminii”, diferând între ei doar prin probabilitatea de a absorbi fotoni de lungimi de undă diferite.
Când o moleculă de rodopsina absoarbe 1 cuantum de lumină, are loc o modificare a configurației spațiale a unei molecule de retină - fotoizomerizarea, care constă în transformarea formei de 11 cm a retinei într-o transformare completă (Fig. 3.1.4b) .
Fotoizomerizarea este însoțită de un ciclu de reacții fotochimice, pentru a căror decodare G. Wald a fost distins cu Premiul Nobel în 1930. Produșii finali ai fotoizomerizării sunt complet traxretinale și opsina. Toate reacțiile intermediare până la etapa metarhodopsin-II sunt reversibile: produsele acestor reacții pot fi convertite înapoi în rodopsina prin izomerizare inversă gran-retiniană.
Aceasta înseamnă că nu apar modificări conformaționale semnificative în molecula de pigment înainte de metarhodopsin-II. În stadiul de metarhodopsin-II, retina este detașată din partea proteică a moleculei. Dintre toate etapele izomerizării, doar prima - transformarea rodopsinei în prelumirodopsină (bathorhodopsin) - are nevoie de lumină. Toate celelalte reacții au loc în întuneric. Fotoizomerizarea are loc în mod similar în pigmenții conici.
Astfel, fotoizomerizarea este un mecanism molecular de declanșare (de pornire) al fotorecepției, care constă în modificarea configurației spațiale a moleculei de pigment vizual. Acesta din urmă clarifică de ce natura a ales carotenoizii ca pigment vizual: prezența legăturilor duble în retină determină posibilitatea izomerizării acesteia și; reactivitatea sa ridicată. În jurul fiecărei legături duble, părți ale moleculei se pot roti și îndoi în spațiu. Această formă este cea mai puțin stabilă și cea mai sensibilă la temperatură și lumină. Găsit în natură tipuri diferite retiniană - 7, 9, 11, 13-cis-forme, 9,13-dicis-form, dar în pigmenții vizuali retina este întotdeauna în 11-cis-form.
Resinteza pigmentului în fotoreceptori are loc continuu la lumină și întuneric cu participarea enzimei retinen izomerazei și constă în izomerizarea inversă a retinolului din transformare la forma 11-cis. În conuri, această enzimă este localizată în receptorii înșiși, în bastonașe, în epiteliul pigmentar, unde trans-retinolul trece după desprinderea de opsină și unde este izomerizat în forma 11-cis. În acest caz, vitamina A retiniană este refolosită, iar proteina este sintetizată în ribozomii segmentului interior.
Se presupune că recombinarea 11-cis-retinalului cu opsina are loc spontan. Epiteliul pigmentar conține aproximativ 11% vitamina A (retinol) din 13% în întregul ochi. Vitamina A se formează în ficat prin ruperea în două părți a lanțului carotenoidului conținut de alimente și prin adăugarea de apă.
Vitamina A este transportată de o proteină specială care leagă retinolul în epiteliul pigmentar, unde este oxidată la retinină. Cu o deficiență de vitamina A, apare așa-numita „orbire nocturnă” - o scădere a sensibilității absolute la lumină, care afectează în special vederea la amurg, transformându-se în orbire permanentă din cauza distrugerii unei proteine care este instabilă în stare liberă. Prin urmare, cu o boală de „orbire nocturnă” se recomandă consumul de morcovi care conțin provitamina A - beta-caroten.
Fiecare fotoreceptor conține un singur fotopigment, caracterizat printr-unul sau altul spectru de absorbție. În acest sens, sunt izolate conuri de lungime de undă scurtă, medie și lungă, care conțin pigmenți la primate cu maxime de absorbție, respectiv, la 445 nm, 535 nm și, respectiv, 570 nm. La peste de apa dulce conul de lungime de undă lungă conține un pigment cu un maxim de absorbție la 620 nm, care se corelează cu forma distribuției spectrale a luminii în habitatul lor.
Aceste conuri sunt uneori denumite receptori sensibili la albastru, verde și roșu, urmând o tradiție care datează din ipoteza cu trei componente Young-Helmholtz. Dar încă din a doua jumătate a secolului al XX-lea, când s-a constatat că niciun fotoreceptor nu realizează singur analiză spectrală, ci reacționează doar la numărul de fotoni absorbiți, indiferent de lungimea de undă a radiației, aceste nume de culoare ale receptorilor sunt folosite doar într-un mod metaforic. sens.
Clasificarea tijelor, de exemplu, la o broască, în „verde” și „roșu” în unele cazuri se bazează nu pe sensibilitatea spectrală a pigmentului conținut în acestea, ci pe culoarea corpurilor receptori în lumina transmisă. Astfel, tijele „verzi” conțin un pigment cu o absorbție maximă la 432 nm, prin urmare, atunci când sunt iluminate cu lumină albă, ele absorb radiația de unde scurte, trecând toate celelalte lungimi de undă și arată verzui când sunt controlate vizual.
În retina peștilor, s-a observat o relație între lungimea conului și tipul spectral al pigmentului conținut în acesta: cele mai scurte dintre ele conțin un pigment cu undă scurtă, iar cele mai lungi un pigment cu undă lungă. Eberle a sugerat că acesta ar putea fi unul dintre mecanismele de combatere a aberației cromatice. Datorită aberației cromatice, imaginea clară a radiațiilor cu undă medie este situată mai adânc (mai departe de lentilă) decât imaginea clară a radiației cu unde scurte, iar focalizarea radiației cu undă lungă este situată și mai departe.
Este suficient să aranjați receptorii cu lungime de undă scurtă, medie și lungă la nivelurile adecvate, astfel încât toate părțile radiației spectrale să fie focalizate la fel de clar asupra receptorilor. Acest lucru, poate, explică faptul că în retina peștilor, conurile simple cu segmente scurte conțin un pigment cu lungime de undă scurtă, în timp ce într-un con dublu, segmentul cel mai lung conține un pigment cu lungime de undă lungă, iar cel mai scurt conține un pigment scurt. sau pigment cu lungime de undă medie. Conurile duble cu segmente egale în lungime conțin același pigment.
ωπς - ochi) - denumirea comună mai mulți pigmenți vizuali ai oamenilor și a unor mamifere. Acești pigmenți sunt alcătuiți dintr-o moleculă proteică asociată cu cromoforul retinian. Conținut în conurile retinei ochiului și oferă viziunea colorată, spre deosebire de un alt pigment vizual - rodopsina, responsabil pentru vederea crepusculară.S-a stabilit că compoziția pigmenților vizuali include opsine. Opsine diferite diferă în compoziția lor de aminoacizi și absorb lumina la lungimi de undă ușor diferite, cum ar fi moleculele legate de retină.
Existența unui pigment conic a fost descoperită pentru prima dată (indirect) de George Wald, care i-a dat numele. iodopsină. . În 1967 i s-a acordat Premiul Nobel pentru Fiziologie sau Medicină pentru aceste lucrări.
Chiar și Helmholtz credea că în „aparatul terminal al fibrelor nervoase optice” (fotoreceptorii retinei) ar trebui să existe trei tipuri de substanțe descompuse fotochimic, care au sensibilitate diferită în ceea ce privește părți diferite spectru. .
Conform teoriei cu trei componente a vederii care predomină în știința modernă, se crede că există trei tipuri de acest pigment și, respectiv, retina conține trei tipuri de conuri care sunt sensibile la lumina albastră, verde și roșie. În consecință, iodopsina, situată în conurile ochiului, constă din trei pigmenți - chlorolab, erythrolab și cyanolab; prima dintre ele absoarbe razele corespunzătoare galben-verde (banda de absorbție 450-630 nm), a doua - galben-roșu (500-700 nm) și a treia - albastru-verde (500-700 nm) partea vizibilă. spectru.
Tipuri de pigmenți vizuali
Primele încercări de a găsi trei pigmenți și, așa cum era de așteptat, trei tipuri de conuri (pe baza ipotezei vederii cu trei componente conform căreia fiecare con conține un singur pigment) au fost efectuate de Rushton, care a perfecționat tehnica densitometriei pentru in vivo. măsurarea coeficienților de absorbție a luminii cu lungimi de undă diferite – în stratul fotoreceptorilor retinieni. S-a demonstrat că anomaliilor de culoare le lipsește unul dintre pigmenții prezenți la persoanele cu vedere normală: „erythrolab” (maximum aproximativ 585 nm.) în protanop și „chlorolab” (maximum aproximativ 540 nm.) în deuteranop.
Trebuie remarcat faptul că atunci când se utilizează termenul „receptor”, în unele cazuri, întregul set de fotoreceptori cu aceeași sensibilitate spectrală este considerat ca unul receptor; în alte cazuri, întrebarea este dacă fiecare con foveal conține trei receptor sau doar unul. În același timp, nu este încălcată strictețea conceptului de „receptor”, care în acest caz nu include caracteristici morfologice specifice.
Următorul pas în această direcție a fost studiul fotopigmenților conținute în conurile individuale de oameni și macac. Dimensiunile conurilor foveale sunt prea mici pentru a face obiectul studiului, iar toate datele obținute se referă doar la conurile parafoveale. Fiecare con, cel puțin cel extrafoveal, pare să conțină doar unul dintre pigmenți, sau predominant unul dintre ei.
Metode moderne de studiere a fotopigmenților conici
Densitometria celulelor retiniene
→ densitometrie, pot fi următoarele:
Dacă primele încercări de a găsi trei pigmenți și, așa cum era de așteptat, trei tipuri de conuri au fost efectuate de Rushton, care a perfecționat tehnica Densitometrie pentru a măsura coeficienții de absorbție a luminii cu lungimi de undă diferite - în stratul fotoreceptorilor retinieni și s-a demonstrat că anomaliilor de culoare le lipsește unul dintre pigmenții pe care îi au persoanele cu vedere normală: „erythrolab” (maximum aproximativ 585 nm.) în protanop și „ chlorolab" (maximum aproximativ 540 nm.) - în deuteranop, acum cu ajutorul unor metode speciale de distometrie folosind densitometre moderne, oamenii de știință reușesc să determine funcționarea conurilor și tijelor în stare normală de activitate și să le diagnosticheze bolile.
Vezi si
Surse
Literatură
- Khokhlova T.V. Idei moderne despre viziunea mamiferelor // Journal of General Biology. Volumul 73, 2012. Nr 6, noiembrie-decembrie. Pagină 418-434.


