Vizuális pigmentek. A bőr epidermiszének kezdetleges zónája rétegekből áll. Nappali és éjszakai látás
A vizuális pigmentek a külső szegmensek membránjaiban koncentrálódnak. Minden rúd körülbelül 10 8 pigmentmolekulát tartalmaz. Több száz diszkrét korongba rendeződnek (körülbelül 750 egy majomrúdban), amelyek nem kapcsolódnak a külső membránhoz. A kúpokban a pigment speciális pigment redőkben található, amelyek a fotoreceptor külső sejtmembránjának folytatásai. A pigmentmolekulák az összes lemezfehérje körülbelül 80%-át teszik ki. A vizuális pigmentek a külső szegmens membránjaiban olyan sűrűn vannak elhelyezve, hogy a pálcában lévő két vizuális pigmentmolekula közötti távolság nem haladja meg a 10 nm-t. Ez a sűrű tömítés növeli annak valószínűségét, hogy a fotoreceptor sejtrétegen áthaladó fény fotonja befogható lesz. Felmerül a következő kérdés: hogyan keletkeznek jelek, amikor a fényt a vizuális pigmentek elnyelik?
Fényelnyelés vizuális pigmentek általPszichofiziológiai, biokémiai és molekuláris technikákkal tanulmányozták azokat az eseményeket, amelyek akkor következnek be, amikor a rúdpigment, a rodopszin elnyeli a fényt. A vizuális pigmentmolekula két komponensből áll: egy opszin nevű fehérjéből és egy kromoforból, egy 11-cisz-A-vitamin aldehidből, amelyet retinának neveznek (1. ábra). Tisztázni kell, hogy a kromofor olyan kémiai csoportot tartalmaz, amely színt ad a vegyületnek. A pigmentek abszorpciós képességének mennyiségi jellemzőit spektrofotometriával vizsgáltuk. Amikor a rodopszint, a rudak vizuális pigmentjét különböző hullámhosszú fénnyel világították meg, a kékeszöld, körülbelül 500 nm hullámhosszú fény nyelődött el a legjobban. Hasonló eredményt kaptunk, amikor egy külön rudat mikroszkóp alatt különböző hullámhosszú fénysugarakkal világítottak meg. Beazonosították érdekes függőség a rodopszin abszorpciós spektruma és az alkonyi fény érzékelése között. Embereken végzett kvantitatív pszichofizikai vizsgálatok kimutatták, hogy a körülbelül 500 nm hullámhosszú kékes-zöld nappali fény optimális a szürkületi fény érzékeléséhez sötétben. Napközben, amikor a rudak inaktívak és csak kúpokat használunk, a kúpok abszorpciós spektrumának megfelelő vörös színre vagyunk a legérzékenyebbek (erről később lesz szó).
Amikor a rodopszin elnyel egy fotont, a retina fotoizomerizálódik, és a 11-cisz-ből transz-konfigurációba kerül. Ez az átmenet nagyon gyorsan megtörténik: körülbelül 10-12 másodperc alatt. Ezt követően a pigment fehérje része is egy sor átalakuláson megy keresztül, számos köztes termék keletkezésével. A fehérjerész egyik konformációja, a metarodopszin II a legfontosabb a jelátvitelhez (erről a fejezet későbbi részében fogunk beszélni). A 2. ábra az aktív rodopszin fehérítésének és regenerációjának eseménysorát mutatja. A metarodopszin II 1 ms alatt képződik. A pigment regenerációja a bomlás után lassan, néhány percen belül megy végbe; Ez megköveteli a retina transzportját a fotoreceptoroktól a pigment epitéliumhoz.
A rodopszin szerkezeteMolekuláris szinten az opszin fehérje 348 aminosavból áll, amelyek 7 hidrofób zónát alkotnak, amelyek mindegyike 20-25 aminosavból áll, és 7 transzmembrán hélixet alkot. A molekula N-vége az extracelluláris térben (azaz a rúdkorong belsejében), a C-vége pedig a citoplazmában található.
Sötétben a 11-cisz-retinál szorosan kötődik az opszin fehérjéhez. A fotonbefogás az összes cisz-retina izomerizációját eredményezi tron retinává. Ebben az esetben az opszin minden tronból és retinából álló komplex gyorsan metarodo psin II-vé alakul, amely opszinná és minden tronból álló retinává disszociál. A rodopszin regenerációja a fotoreceptorok és a pigmenthámsejtek kölcsönhatásától függ. A Metarhodopsin II bekapcsolja és aktív állapotban tartja a másodlagos hírvivő rendszert.
A retina a hetedik transzmembrán szegmensben található lizin maradékon keresztül kapcsolódik az opszinhoz. Az Opsin a 7 transzmembrán domént tartalmazó fehérjék családjába tartozik, amely metabotróp neurotranszmitter receptorokat, például adrenerg és muszkarin receptorokat is tartalmaz. A rodopszinhoz hasonlóan ezek a receptorok a G-fehérje aktiválásával továbbítják a jeleket a másodlagos hírvivőknek. A rodopszin meglepően stabil a sötétben. Bayor számításai szerint egy rodopszin molekula spontán termikus izomerizációjához körülbelül 3000 évre van szükség, vagyis 10 23-mal többet, mint a fotoizomerizációhoz.
Vizuális pigmentek
Vizuális pigmentek
A rodopszin szerkezete
Kúpok és színlátás
Színvakság
Irodalom
Vizuális pigmentek
A vizuális pigmentek a külső szegmensek membránjaiban koncentrálódnak. Minden rúd körülbelül 10 8 pigmentmolekulát tartalmaz. Több száz diszkrét korongba rendeződnek (körülbelül 750 egy majomrúdban), amelyek nem kapcsolódnak a külső membránhoz. A kúpokban a pigment speciális pigment redőkben található, amelyek a fotoreceptor külső sejtmembránjának folytatásai. A pigmentmolekulák az összes lemezfehérje körülbelül 80%-át teszik ki. A vizuális pigmentek a külső szegmens membránjaiban olyan sűrűn vannak elhelyezve, hogy a pálcában lévő két vizuális pigmentmolekula közötti távolság nem haladja meg a 10 nm-t. Ez a sűrű tömítés növeli annak valószínűségét, hogy a fotoreceptor sejtrétegen áthaladó fény fotonja befogható lesz. Felmerül a következő kérdés: hogyan keletkeznek jelek, amikor a fényt a vizuális pigmentek elnyelik?
Fényelnyelés vizuális pigmentek által
Pszichofiziológiai, biokémiai és molekuláris technikákkal tanulmányozták azokat az eseményeket, amelyek akkor következnek be, amikor a rúdpigment, a rodopszin elnyeli a fényt. A vizuális pigmentmolekula két komponensből áll: egy opszin nevű fehérjéből és egy kromoforból, egy 11-cisz-A-vitamin aldehidből, amelyet retinának neveznek (1. ábra). Tisztázni kell, hogy a kromofor olyan kémiai csoportot tartalmaz, amely színt ad a vegyületnek. A pigmentek abszorpciós képességének mennyiségi jellemzőit spektrofotometriával vizsgáltuk. Amikor a rodopszint, a rudak vizuális pigmentjét különböző hullámhosszú fénnyel világították meg, a kékeszöld, körülbelül 500 nm hullámhosszú fény nyelődött el a legjobban. Hasonló eredményt kaptunk, amikor egy külön rudat mikroszkóp alatt különböző hullámhosszú fénysugarakkal világítottak meg. Érdekes összefüggést fedeztek fel a rodopszin abszorpciós spektruma és az alkonyi fény észlelése között. Embereken végzett kvantitatív pszichofizikai vizsgálatok kimutatták, hogy a körülbelül 500 nm hullámhosszú kékes-zöld nappali fény optimális a szürkületi fény érzékeléséhez sötétben. Napközben, amikor a rudak inaktívak és csak kúpokat használunk, a kúpok abszorpciós spektrumának megfelelő vörös színre vagyunk a legérzékenyebbek (erről később lesz szó).
Amikor a rodopszin elnyel egy fotont, a retina fotoizomerizálódik, és a 11-cisz-ből transz-konfigurációba kerül. Ez az átmenet nagyon gyorsan megtörténik: körülbelül 10-12 másodperc alatt. Ezt követően a pigment fehérje része is egy sor átalakuláson megy keresztül, számos köztes termék keletkezésével. A fehérjerész egyik konformációja, a metarodopszin II a legfontosabb a jelátvitelhez (erről a fejezet későbbi részében fogunk beszélni). A 2. ábra az aktív rodopszin fehérítésének és regenerációjának eseménysorát mutatja. A metarodopszin II 1 ms alatt képződik. A pigment regenerációja a bomlás után lassan, néhány percen belül megy végbe; Ez megköveteli a retina transzportját a fotoreceptoroktól a pigment epitéliumhoz.
A rodopszin szerkezete
Molekuláris szinten az opszin fehérje 348 aminosavból áll, amelyek 7 hidrofób zónát alkotnak, amelyek mindegyike 20-25 aminosavból áll, és 7 transzmembrán hélixet alkot. A molekula N-vége az extracelluláris térben (azaz a rúdkorong belsejében), a C-vége pedig a citoplazmában található.
1. ábra. A fotoreceptor membránba ágyazott gerinces rodopszin szerkezete. A hélix kissé el van forgatva, hogy megmutassa a retina elhelyezkedését (feketével jelölve). C - C-terminális, N - N-terminális.
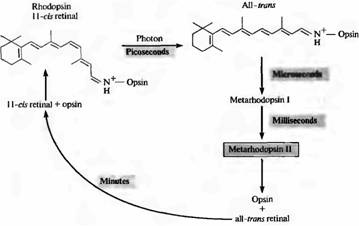
2. ábra. A rodopszin elhalványulása fényben. Sötétben a 11-cisz-retinál szorosan kötődik az opszin fehérjéhez. A fotonbefogás az összes cisz-retina izomerizációját eredményezi tron retinává. Ebben az esetben az opszin minden tronból és retinából álló komplex gyorsan metarodo psin II-vé alakul, amely opszinná és minden tronból álló retinává disszociál. A rodopszin regenerációja a fotoreceptorok és a pigmenthámsejtek kölcsönhatásától függ. A Metarhodopsin II bekapcsolja és aktív állapotban tartja a másodlagos hírvivő rendszert.
A retina a hetedik transzmembrán szegmensben található lizin maradékon keresztül kapcsolódik az opszinhoz. Az Opsin a 7 transzmembrán domént tartalmazó fehérjék családjába tartozik, amely metabotróp neurotranszmitter receptorokat, például adrenerg és muszkarin receptorokat is tartalmaz. A rodopszinhoz hasonlóan ezek a receptorok a G-fehérje aktiválásával továbbítják a jeleket a másodlagos hírvivőknek. A rodopszin meglepően stabil a sötétben. Bayor számításai szerint egy rodopszin molekula spontán termikus izomerizációjához körülbelül 3000 évre van szükség, vagyis 10 23-mal többet, mint a fotoizomerizációhoz.
Kúpok és színlátás
Young és Helmholtz bámulatos kutatásai és kísérletei a 19. században felhívták a figyelmet nagyon fontos kérdés a színlátásról, és maguk a tudósok is világos és pontos magyarázatot adtak erre a jelenségre. Következtetésük három létezéséről különféle típusok A színes fotoreceptorok kiállták az idő próbáját, és ezt követően molekuláris szinten is megerősítették. Ismét idézhetjük Helmholtzot, aki a fény és a hang, a szín és a hangtónus érzékelését hasonlította össze. Gondolatának tisztaságát, erejét és szépségét irigyelni lehet, különösen a 19. században elterjedt zavaros vitalisztikus fogalmakhoz képest:
A színtónusok minden különbsége a három alapszín – vörös, zöld és lila – eltérő arányú kombinációjától függ... Ahogy a napfény és annak melegének érzékelése is attól függ, hogy a napsugarak elérik-e a idegek, amelyek látásreceptorokból vagy hőérzékenységi receptorokból származnak. Ahogyan Young hipotézisében javasolta, a különböző színek érzékelésének különbsége egyszerűen attól függ, hogy a 3 típusú fotoreceptor közül melyiket aktiválja jobban az adott fény. Amikor mindhárom típus benne van egyaránt izgatott, fehér lesz...
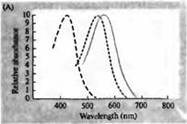
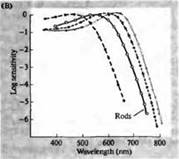
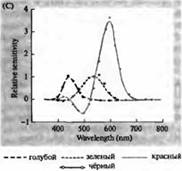
Rizs. 3. Az emberi fotoreceptorok érzékenységi spektrumai és különféle vizuális pigmentek. (A) Három színes vizuális pigment érzékenységi spektruma görbéi, amelyek abszorpciós csúcsokat mutatnak a kéknek, zöldnek és vörösnek megfelelő hullámhosszon. (B) A kúpok kékre, zöldre és vörösre, valamint (feketével látható) rudak érzékenységi spektruma makákóknál. A válaszokat szívóelektródákkal rögzítettük, átlagoltuk és normalizáltuk. A rúdspektrum görbéket a vizuális pigmentek emberen végzett vizsgálataiból nyerték. (C) A majom és az ember kúpspektrumának összehasonlítása színérzékenységi teszttel. A folytonos görbe egy kísérletet mutat be emberek színérzékenységének meghatározására különböző hullámhosszúságú fény bemutatásával. A szaggatott vonal az egyes kúpok áramainak rögzítéséből származó előrejelzett eredményeket mutatja, miután korrigáltuk a lencsében és a pigmentekben a külső szegmens felé vezető fényelnyelést. A két kísérlet eredményei között meglepően magas az egyezés.
Ha egyszerre két különböző színű fénysugarat vetítünk egy fehér képernyőre... csak egy színt látunk, amely többé-kevésbé különbözik mindkét színtől. Jobban megérthetjük azt a figyelemre méltó tényt, hogy a külső fény összetételében lévő összes árnyalatot képesek vagyunk érzékelni a három alapszín keverékével, ha összehasonlítjuk a száraz szemet... Hang esetében... halljuk a hosszabb hullámok alacsony hangokként, a rövidebbek pedig - mint magasak és átütőek, ráadásul a fül sok hanghullámot képes egyszerre megfogni, pl. sok jegyzet. Azonban ebben az esetben nem olvadnak össze egyetlen összetett akkordba, mint ahogy a különböző színek... egyesülnek egy összetett színbe. A szem nem tudja megkülönböztetni, ha cseréljük narancsszín pirosra vagy sárgára; de ha a C és E hangot egyszerre halljuk megszólalni, akkor egy ilyen hang nem tűnik D hangnak. Ha a fül ugyanúgy érzékeli a zenei hangokat, mint a szem a színeket, akkor minden akkordot három állandó hang kombinációjával ábrázolhatnánk, egy nagyon mély, egy nagyon magas és egy közöttük lévő hang kombinációjával, amelyek minden lehetséges zenei hatást csak a hangok változásai révén okoznak. ennek a három hangnak a relatív hangossága... Azonban a színek zökkenőmentes átmenetét láthatjuk egyikről a másikra végtelen számú árnyalaton és átmeneten keresztül... A színek érzékelésének módja... elsősorban attól függ a mi felépítésünkről idegrendszer. El kell ismerni, hogy jelenleg sem az emberek, sem a négylábúak esetében nem írták le a színérzékelés elméletének megerősítésének anatómiai alapját.
Ezeket a pontos és előrelátó előrejelzéseket különböző megfigyelések sorozata erősítette meg. Spektrofotometriával Wald, Brauk, McNicol és Dartnall és munkatársai háromféle, különböző pigmenttel rendelkező kúp jelenlétét mutatták ki az emberi retinában. Baylornak és kollégáinak sikerült elterelnie az áramokat a majmok és az emberek kúpjairól is. Megállapították, hogy a három kúppopuláció különálló, de egymást átfedő érzékenységi tartományokkal rendelkezik a spektrum kék, zöld és vörös részéhez. Az izgató elektromos jelek optimális hullámhossza pontosan egybeesett a vizuális pigmentek fényelnyelési csúcsaival, amelyeket spektrofotometriával és a szem színspektrumra való érzékenységét mérő pszichofizikai kísérletekkel állapítottak meg. Végül Natais klónozta és szekvenálta az opszin pigmentet kódoló géneket háromféle kúpban, amelyek érzékenyek a vörös, zöld és kék spektrumra.
Hogyan lehet az, hogy a különböző vizuális pigmentek molekulái egy bizonyos hullámhosszú fényt előszeretettel képesek befogni? Kiderült, hogy a rodopszin a rudak vizuális pigmentje, és a kúpok mindhárom vizuális pigmentje ugyanazt a kromofort, a 11-cisz-retinált tartalmazza. A pigment fehérje részének aminosavszekvenciája azonban különbözik egymástól. A mindössze néhány aminosav különbsége megmagyarázza a spektrum iránti eltérő érzékenységüket.
Színvakság
Bár a fotoreceptorok egyik típusa önmagában nem képes a színek érzékelésére, a háromféle kúp, amint az az ábrán látható. 4 már képes.
A színfelismeréshez elvileg kétféle, eltérő pigmenttel rendelkező kúp is elegendő lenne, de ebben az esetben számos hullámhossz-kombinációt egyformán érzékelnénk. Hasonló helyzet áll elő, ha egy személy színvakságban szenved. Az ilyen embereknek, amint azt Nathans kimutatta, genetikai hibáik vannak, amelyek az egyik pigment hiányához vezetnek. A tudomány jelenlegi állása szerint nem győzünk csodálkozni azon, hogy a molekuláris mechanizmusok milyen gyönyörűen igazolják Young és Helmholtz briliáns és meglepően pontos gondolkodását.
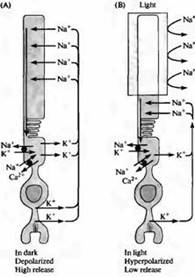
Rizs. 4. "Sötét" áram a rúdban. (A) Sötétben a nátriumionok áthaladnak a rudak külső szegmensében lévő kationcsatornákon, depolarizációt okozva; A kalciumionok is képesek áthaladni ezeken a kationcsatornákon. Az áramhurok áthalad a rúd isthmusán, mivel a káliumáram kifelé halad a membrán belső szegmensében. (B) Amikor a külső szegmens megvilágosodik, a csatornák bezáródnak a cGMP intracelluláris koncentrációjának csökkenése miatt, és a rúd hiperpolarizálódik. A hiperpolarizáció az adó felszabadulásának csökkenéséhez vezet. A nátrium, kálium és kalcium koncentrációját a rúd belsejében speciális szivattyúk és ioncserélők tartják fenn, amelyek a belső szegmens (fekete körök) területén helyezkednek el. A kalcium transzporterek szintén a külső szegmensben találhatók.
Elképzelésük, hogy a színlátás és színvakság alapvető tulajdonságait magukban a fotoreceptorokban kell megtalálni, direkt fiziológiai mérésekkel, valamint a pigmentek genetikai és fehérjeszintű szerkezeti különbségeinek vizsgálatával igazolták, a potenciál nagyságrendileg nagy. -40 mV, ami messze van az E K egyensúlyi káliumpotenciáltól, ami 80 mV.

Rizs. 5. A cGMP szerepe a pálcikák külső szegmensének nátriumcsatornáinak megnyitásában. Az egyes csatornák aktivitását különböző koncentrációjú cGMP-t tartalmazó oldatban elhelyezett, belülről kifelé tartó patch clamp segítségével rögzítettük. A csatorna kinyitása az áram felfelé tereléséhez vezet. A csatornanyitás gyakorisága nagyon alacsony volt a kontrollban, a cGMP hozzáadása az egycsatornás nyitás gyakoriságának növekedéséhez vezetett, ami közvetlenül függött a cGMP koncentrációjától
A sötétben bejövő áramot elsősorban nátriumionok viszik, követve az elektrokémiai gradienst a külső szegmens kationcsatornáin keresztül. A fotoreceptor fény hatására bekövetkező hiperpolarizációját ezeknek a csatornáknak a bezáródása okozza, ami a membránpotenciál E K felé tolódásához vezet.
A fotoreceptor csatornák tulajdonságai
A külső szegmens kationcsatornáinak fiziológiás körülmények között a kalcium/nátrium/kálium ion vezetőképesség aránya 12,5:1,0:0,7, az egycsatornás vezetőképesség pedig körülbelül 0,1 pS21). Mivel a nátrium koncentrációja sokkal magasabb, mint a kalcium koncentrációja, a bejövő áram körülbelül 85%-a nátriumionokból származik. A káliumionok hajtóereje a sejten kívülre irányul. Amikor a kalciumionok áthaladnak a csatornán, ezek az ionok szilárdan kötődnek a pórusfalhoz, és így akadályozzák más ionok áthaladását. Ennek köszönhető, hogy az extracelluláris környezetből a kalcium eltávolítása a kálium- és nátriumionok könnyebb átjutását eredményezi a csatornákon, amelyek vezetőképessége 25 pS-re nő.
Fesenko, Yau, Baylor, Strayer és munkatársai kimutatták, hogy a ciklikus GMP belső jelhordozó szerepét tölti be a lemeztől a membrán felületéig. Ahogy a 4. ábrán látható, a cGMP magas koncentrációja a citoplazmában nyitva tartja a kationcsatornákat. Mivel a cGMP koncentrációja a membrán belső felületéről csökken, a kationcsatornák felnyílása ritka eseménysé válik. Így a fotoreceptorok membránpotenciálja a cGMP citoplazmában lévő koncentrációját tükrözi: minél magasabb a cGMP koncentrációja, annál nagyobb a sejt depolarizációja. A cGMP koncentrációja viszont a beeső fény intenzitásától függ. A növekvő fényintenzitás a cGMP koncentráció csökkenéséhez vezet, és csökkenti a nyitott csatornák arányát. A cGMP hiányában szinte minden csatorna zárva van, és a külső szegmens membrán ellenállása megközelíti a lipid kettősrétegét.
A cGMP-kapuzott csatornák molekuláris szerkezete
A pálcikák külső szegmensének ioncsatornáihoz cDNS-t izoláltunk, és meghatároztuk ezen csatornák alegységeinek aminosavszekvenciáját emberek, szarvasmarhák, egerek és csirkék retinájában. Jelentős DNS-hasonlóságot találtak ezeknél és más cGMP-kapuzott ioncsatornáknál – például a szaglórendszerben
Jelátvitel fotoreceptorokban
Hogyan vezet a rodopszin fotoizomerizációja a membránpotenciál változásához? Sok éven át megértették, hogy valamiféle belső hírvivőre van szükség az elektromos jelek gerjesztéséhez a rudakban és kúpokban. Az egyik oka annak gyanújának, hogy a rúd külső szegmensének régiójában a fotonok abszorpciójával kapcsolatos információkat hordozó segítségével továbbítják az a tény, hogy maga a rodopszin pigment a korong belsejében található, és a jel a citoplazmán keresztül terjed. a külső membrán. A második ok a válaszadás jelentős növekedése volt. Baylor és munkatársai a teknősök fotoreceptorait tanulmányozva kimutatták, hogy a membrán vezetőképessége és a rögzített elektromos jelek csökkenése már csak egy foton elnyelése és a 108 pigmentmolekula egyikének aktiválása esetén is bekövetkezik.
Az események sorozatát, amelyek során egy aktivált fotopigment molekula megváltoztatja a membránpotenciált, a rudak és kúpok külső szegmenseinek tanulmányozásával sikerült tisztázni, kételektródos potenciálleszorító és molekuláris biológiai technikákkal. A fényfoton abszorpciójától az elektromos jelig terjedő jelátviteli diagramot az ábra mutatja. 3.
Sötétben állandó bejövő „sötét” áram folyik a rudak és kúpok külső szegmenseiben. Membránrendszerük következtében. Ezen csatornák membránrégiói szerkezeti hasonlóságot mutatnak más kation-szelektív csatornákkal, különösen az S4 régióban és az ionpórust alkotó régióban. A fotoreceptor ioncsatornák legalább 2 különböző fehérje alegységből álló tetramerek, amelyek molekulatömege 63, illetve 240 kDa.
Az intracelluláris nukleotidkötő helyek az alegységek karboxilvégének közelében helyezkednek el. Ezen alegységek petesejtekben történő expressziója olyan kationcsatornákat eredményez, amelyek hasonló tulajdonságokkal rendelkeznek, mint a pálcikák külső szegmenseiben: a cGMP aktiválja őket, és rendelkeznek a várt vezetőképesség-permeabilitás aránnyal.

Rizs. 6. A G-protein aktiválódásának mechanizmusa fotopigment molekula aktiválódása során. A G-protein transzducin megköti a GTP-t metarodopszin II jelenlétében, ami a foszfodiészteráz aktiválásához vezet, ami viszont hidrolizálja a cGMP-t. A cGMP-koncentráció csökkenésével a nátriumcsatornák bezáródnak.
A ciklikus GMP metabolikus kaszkád
A cGMP-koncentráció csökkenéséhez és az ioncsatornák ezt követő bezárásához vezető események láncolata az 5. ábrán látható. Az intracelluláris cGMP-koncentráció csökkenését a fény okozza, ami a fotopigment lebomlásának köztiterméke, a metarodopszin II képződéséhez vezet. A metharhodopszin II viszont a G-protein transzducinjára hat, amely 3 polipeptid láncból áll
A metarodopszin II és a transzducin kölcsönhatása a G-proteinhez kötött molekula GDP GTP-vel való helyettesítéséhez vezet. Ez aktiválja a G fehérje alegységet, amely elválik az alegységektől, és viszont aktiválja a membránrégióban található foszfodiészterázt: egy enzimet, amely hidrolizálja a cGMP-t. A cGMP koncentrációja csökken, kevesebb a nyitott ioncsatorna, és a rúd hiperpolarizálódik. A kaszkádot az aktív metarodopszin II C-terminálisának foszforilációja szakítja meg. A cGMP kulcsszerepét a kationcsatornák állapotának szabályozásában biokémiai kísérletek igazolták. A fotoreceptorok megvilágítása 20%-kal csökkentheti a cGMP-szintet a sejtben.
Gerinces receptorok, amelyek fény hatására depolarizálódnak
Érdekes kivétel a fentebb ismertetett fotorecepciós mechanizmus alól néhány gerinces receptor. A gyíkoknak van egy harmadik szeme a fejük tetején. Kis „kúpokat” tartalmaz, amelyek képesek a fő (oldalsó) szem által észlelthez hasonló képet érzékelni. Ezek a fotoreceptorok azonban figyelemre méltóak abban, hogy megvilágítás hatására depolarizálódnak. A nukleotid-kapu csatornák itt hasonló szerkezetűek és funkciójúak, mint más gerincesek fotoreceptorai, egy kivétellel: a fotoreceptor és a G-fehérje aktiválása a cGMP-koncentráció növekedéséhez vezet. Ennek eredményeként a külső szegmens csatornái megnyílnak, és a kationok a cellába irányítva „könnyű” áramot képeznek. Ez a foszfodiészteráz aktivitás gátlása miatt következik be sötétben. Ennek eredményeként a következő eseménysor következik be a gyíkszemben: fény - [cGMP] - a külső szegmens kationcsatornáinak felnyílása - depolarizáció.
Jelerősítés a cGMP kaszkádban
A kétlépcsős cGMP-kaszkád jelentős növekedést biztosít a kezdeti jelben, ami megmagyarázza a rudak kivételes fényérzékenységét. Először is, egy molekula aktív metarodopszin II katalizálja sok GTP molekula hozzáadását a GDP helyett, és így több száz G-fehérje alegységet szabadít fel. Másodszor, minden alegység aktivál egy foszfodiészteráz molekulát a korongban, amely képes lebontani a citoplazmában található hatalmas számú cGMP molekulát, és így nagyszámú csatorna bezárásához vezet.
Jelek válaszként egyetlen fénykvantumra
Bizonyíték arra, hogy egyetlen fénykvantum képes előidézni a fény érzékelését nagyszámú kérdéseket. Mekkora ez az egyetlen válasz? Hogyan választják el ezt a jelet a zajszinttől? És hogyan továbbítják ezeket az információkat megbízhatóan a retináról a magasabb vizuális központokba? Az egyes fénykvantumokra adott jelek mérésére Baylor és munkatársai a varangyok, majmok és emberek retinájában lévő egyes rudak áramlatait rögzítették. Ezek a kísérletek egyedülálló példát mutatnak egy olyan kísérletre, amely megmutatja, hogyan nehéz folyamat, mint a halvány fényvillanások érzékelése, összefüggésbe hozható az egyes molekulák szintjén bekövetkező változásokkal.
A retinatöredék állati vagy holttestből származó anyagból történő izolálására szolgáló eljárást ezekhez a kísérletekhez sötétben kell végrehajtani. Az áramerősség méréséhez a rúd külső szegmensét egy vékony pipettába szívják. Ahogy az várható volt, ezek a kísérletek azt mutatják, hogy sötétben az áram folyamatosan a külső szegmensbe folyik. A fényvillanások a külső szegmens csatornáinak bezárásához vezetnek, ami a "sötét" áram csökkenését okozza. Az áram amplitúdója kicsi és arányos az elnyelt kvantumok számával. A vaku néha egyetlen választ ad, néha kettős választ, néha pedig egyáltalán nem.
A majomrudakban az áram csökkenése egy foton elnyelésére körülbelül 0,5 pA. Ez körülbelül 300 csatorna lezárásának felel meg, i.e. az összes csatorna 3-5%-a sötétben nyílik meg. Ez az anyagcsere jelének jelentős növekedése miatt érhető el cGMP kaszkád. Ezenkívül a vizuális pigmentek korábban említett rendkívüli stabilitása miatt a véletlenszerű izomerizáció és a hamis csatornazáródás nagyon ritka események. Ez azt eredményezi, hogy az egyes fénykvantumok hatásai kiemelkednek a nagyon alacsony állandó zaj háttérben. Bebizonyosodott, hogy a fotoreceptorok közötti réscsatlakozásokon keresztül történő elektromos csatolás további simító hatást biztosít, amely csökkenti a háttérzajt, és javítja a rúdválaszok jel-zaj arányát az egyes kvantumokra.
Irodalom
1. Finn, J. T., Grunwald, M. E. és Yau, K-W. 1996. Ciklikus nukleotid-kapuzott ioncsatornák: Változatos funkciókkal rendelkező, kiterjedt család. Annu. Fordulat. Physiol.58, 395-426.
2. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y., Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S. és Okada, M. 1998. Glutamate receptorok: agyműködés és jelátvitel.
VIZUÁLIS PIGMENTEK(lat. pigmentum festék) - a retina fotoreceptorainak fényérzékeny pigmentjei. 3. A fényimpulzus energiáját érzékelve összetett fotokémiai cikluson mennek keresztül. átalakulások, amelyek eredményeként a retina különálló, 3. p.-ot tartalmazó vizuális receptora (kúp vagy rúd) gerjesztett állapotba kerül, és a kapott információt a látóideg mentén továbbítja a központi idegrendszer felé. n. Val vel. A retina vizuális sejtjei fotoreceptor membránjának fő szerkezeti és funkcionális részeként a 3. p. kulcsszerep a látás mechanizmusaiban (lásd).
A vizuális pigmentek elnevezése és szerkezete. A gerinces és gerinctelen állatok összes vizsgált 3. pontja az opszin vízben oldhatatlan membránfehérje és a kapcsolódó kromofor (retina) komplexei. A retinál vagy az A-vitamin-aldehid két formában létezhet - a retina1 és a retina2 formájában.
A kromofor természete szerint a 3. p. két osztályba sorolható - a retinális1-et tartalmazó rodopsinek (lásd.) és a retinális2-t tartalmazó porfiropzinok. A rodopszinok minden szárazföldi és tengeri állat szemének retinájában, a porfiropzinok az édesvízi állatok szemének retinájában találhatók. Egyes halaknál és kétéltűeknél 3. olyan tételeket találtak, amelyek egyidejűleg tartalmaznak retinát! és retina. Vannak kísérletek a 3. p. osztályozására a retina rúdjaira vagy kúpjaira jellemző opszinok különbségei alapján. Például a rodopszin a retina1 és a rúd opszin komplexe, a jodopszin a retina1 és a kúpos opszin komplexe, a porfiropzin a retina2 komplexe rúd-opzinnal, a retina-kúp opszin komplex cianopzint képez. A 3. o.-t azonban rendkívül nehéz opszinok alapján besorolni, mivel legalább öt különböző opszin létezik.
Az összes ismert 3. p. közül a bikák, békák és tintahalak szeméből izolált rodopszinokat tanulmányozták a legteljesebben. Azt mondják tömeg (tömeg) körülbelül 30-40 ezer, minden molekula kb. 400 aminosav és egy kromofor. Ezenkívül a 3. p. molekula egy oligoszacharid láncot tartalmaz: 3 glükózamin gyök, 2 mannóz, 1 galaktóz. A lipidek (főleg foszfolipidek) erős komplexet alkotnak a 3.p molekulával. A lipidek nélküli 3.p, miközben megtartják alapvető spektrális tulajdonságaikat (lásd Spektrális analízis), számos funkcionálisan fontos tulajdonságot elveszítenek, például a helyreállítási képességüket.
A tiszta retina rendelkezik sárga, abszorpciós spektrumának maximuma a 370 nm tartományban van. Az Opsin színtelen, az abszorpciós maximum az ultraibolya tartományban van (kb. 280 nm). A rodopszin molekula színe vöröses-rózsaszín, maximális abszorpciós spektruma kb. 500 nm. A komplex képződése során bekövetkező ilyen erős spektrális eltolódás oka (370-ről 500 nm-re - az úgynevezett batokróm eltolódás) még nem kapott egyértelmű magyarázatot.
A rodopsinek és porfiropzinok abszorpciós spektruma maximumai a látható spektrum meglehetősen széles tartományát fedik le - rodopsinek esetében 433-562 nm, porfiropzinok esetében pedig 510-543 nm. Ha a béka, ponty és édesvízi teknős ebihal 3. kúpját is porfiropzinnak, azaz 620 nm-en maximális abszorpciós spektrummal rendelkező cianopszinnak tekintjük, akkor ez a régió még szélesebbnek bizonyul. A mikrospektrofotometriás módszerek fejlődése lehetővé tette számos típusú egyetlen fotoreceptor sejt abszorpciós spektrumának meghatározását állatokban és emberekben. A kapott adatok szerint a humán retina 3 p. abszorpciós spektruma a következő: pálcikák 498, kékre, zöldre és vörösre érzékeny kúpok - 440, 535 és 575 nm.
A 3. o. tanulmányozása németül kezdődött. H. Muller kutató, aki 1851-ben leírta, hogy a béka szeméből kivont rózsaszínes-lilás retina hogyan válik sárgássá, majd fehéressé a fényben. 1877-ben F. Boll is leírta ezt a jelenséget, és arra a következtetésre jutott, hogy a retina látósejtjeiben valamiféle vörös fényre érzékeny anyag található, és ennek az anyagnak az elszíneződése a látás mechanizmusával függ össze. A 3. pontok tanulmányozásáért Kuhne (W. Kuhne, 1877) nagy érdeme, akinek sikerült elkülönítenie és részletesen tanulmányoznia a 3. pontokat. Az általa kivont 3.p-t vizuális bíbornak nevezte el, megállapította fehérjetermészetét, megvizsgálta egyes spektrális tulajdonságait és fototranszformációit, és felfedezte, hogy a 3.p sötétben is helyreáll. Amer. nagyban hozzájárult a 3. p. biokémikus és fiziológus J. Wald.
Vizuális pigmentek fototranszformációi. Amikor a 3.p.-t fény éri, fotokémiai ciklus megy végbe bennük. transzformációk, amelyek a retina cisz-transz izomerizációjának elsődleges fotokémiai reakcióján alapulnak (lásd Izomerizmus). Ebben az esetben a kromofor és a fehérje közötti kapcsolat megszakad. A 3. p. átalakulásainak sorrendje a következőképpen mutatható be: rodopszin (a kromofor cisz formában van) -> prelumirodopszin -> lumirrodopszin -> metarhodopsin I -> metarhodopsin II -> opszin fehérje -> kromofor transz formában. Az enzim - retinol-dehidrogenáz - hatására ez utóbbi A-vitaminná alakul, amely a rudak és kúpok külső szegmenseiből a retina pigmentrétegének sejtjeibe kerül. Amikor a szem elsötétül, megtörténik a 3. p regenerációja, a vágás végrehajtásához az A-vitamin cisz-izomerjének jelenléte szükséges, amely a kromofor (A-vitamin aldehid) képződésének kiindulási termékeként szolgál. Ha A-vitamin hiánya vagy hiánya van a szervezetben, akkor a rodopszin képződése megszakadhat, és ennek következtében szürkületi látászavar, ún. éjszakai vakság (lásd Hemeralopia). A rodopszin fototranszformációinak folyamatában a lumirhodopszin metarodopszin I-re való átmenetének szakaszában a receptorsejtben fényes villanás hatására következik be az ún. korai (rövid látenciájú) receptorpotenciál. Ő azonban nem vizuális jel, bár az egyik tesztként szolgálhat a fotoreceptor membránban a 3. p. A funkcionális jelentősége az ún. késői receptorpotenciál, melynek látens periódusa (5-10 ms) összemérhető a metarodopszin II képződési idejével. Feltételezhető, hogy a metarodopszin I metarodopszin II átalakulásának reakciója biztosítja a vizuális jel megjelenését.
Mivel a 3. o.-ok folyamatosan elszíneződnek a fényben, léteznie kell a folyamatos helyreállításuknak. Egyesek rendkívül gyorsak (fotoregeneráció), mások meglehetősen gyorsak (biokémiai, regenerációs, sötét), mások lassúak (3. p. szintézise a látósejt fotoreceptor membránjának állandó megújulása során). A fotoregeneráció fiziol, gerinctelen állatoknál fontos (pl. fejlábúak- tintahal, polip). A biokémiai mechanizmusban A 3. p. regenerációjában gerinceseknél láthatóan fontos szerepe van az izomeráz enzimnek (lásd), amely biztosítja a transz-retinális (vagy transz-A-vitamin) izomerizációját ismét cisz-izomer formává. Egy ilyen enzim létezésére azonban még nem áll rendelkezésre végleges bizonyíték. A 3.p molekula kialakulásának reakciója a retina 11-cisz-izomerje és az opszin jelenlétében könnyen, energiafelhasználás nélkül megy végbe. Felfedezték a fehérített rodopszin azon képességét, hogy foszforilációs reakción menjen keresztül (lásd); Feltételezzük, hogy ez a reakció a látósejt fényadaptációjának mechanizmusának egyik láncszeme.
Bibliográfia: Akkerman Yu. Biofizika, ford. angolból, M., 1964; Willie K. és Det e V. Biology, ford. angol nyelvből, M., 1974, bibliogr.; Konev S. V. és Volotovsky I. D. Bevezetés a molekuláris fotobiológiába, p. 61, Minszk, 1971; Ostrovsky M. A. és Fedorovich I. B. Photo-induced changes in the photoreceptor membrán, a könyvben: Structure and functions of biol, membránok, szerk. A.S. Troshina és mtsai, 1. o. 224, M., 1975, bibliogr.; Érzékszervi rendszerek élettana, szerk. Gershuni G. V., 1. rész, 1. o. 88, L., 1971; Vizuális pigmentek biokémiája és élettana, szerk. szerző: H. Langer, V. a. o., 1973; Érzékélettani kézikönyv, szerk. írta: H. A. R. Jung a. o., v. 7, 1-2. pont, B., 1972.
M. A. Osztrovszkij.


