Vizualni pigmenti. fotorecepcija. Katera od membran zrkla je spredaj prozorna. Lateralno genikulatno telo je struktura
VIZUALNI PIGMENTI(lat. pigmentum paint) - fotosenzitivni pigmenti fotoreceptorjev mrežnice. 3. Predmeti, ki zaznajo energijo svetlobnega impulza, opravijo zapleten fotokemični cikel. transformacije, zaradi katerih ločen vizualni receptor mrežnice, ki vsebuje 3. p (stožec ali palico), preide v vzburjeno stanje in prenaša prejete informacije v centralni živčni sistem vzdolž optičnega živca. n. z. Ker je glavni strukturni in funkcionalni del fotoreceptorske membrane vidnih celic mrežnice, 3. str. ključna vloga v mehanizmih vida (glej).
Nomenklatura in struktura vizualni pigmenti. Vse proučevane 3.p vretenčarjev in nevretenčarjev so kompleksi v vodi netopnega membranskega proteina opsina in pripadajočega kromoforja (retinala). Retinal ali aldehid vitamina A lahko obstaja v dveh oblikah - retinal1 in retinal2.
Glede na naravo kromoforja so 3. str razdeljeni v dva razreda - rodopsine (glej), ki vsebujejo retinal1, in porfiropsine, ki vsebujejo retinal2. Rodopsine najdemo v mrežnici oči vseh kopenskih in morskih živali, porfiropsine najdemo v mrežnici oči sladkovodnih živali. Pri nekaterih ribah in dvoživkah so našli 3. predmete, ki hkrati vsebujejo retinal! in mrežnice. Obstajajo poskusi razvrstitve 3. str na podlagi razlik v opsinih, značilnih za paličice ali stožce mrežnice. Na primer, rodopsin je kompleks retinala1 s paličastim opsinom, jodopsin je kompleks retinala1 s stožčastim opsinom, porfiropsin je kompleks retinala2 s paličnim opsinom, kompleks opsina retinal-stožček tvori cianopsin. Je pa izredno težko razvrstiti 3. p. na podlagi opsinov, saj obstaja vsaj pet različnih opsinov.
Od vseh znanih 3. p., so najbolj v celoti raziskani rodopsini, izolirani iz oči bikov, žab in lignjev. Pravijo teža (masa) je približno 30-40 tisoč, vsaka molekula vsebuje cca. 400 aminokislin in en kromofor. Poleg tega molekula 3. p vključuje oligosaharidno verigo: 3 glukozaminske radikale, 2 manozo, 1 galaktozo. Lipidi (predvsem fosfolipidi) tvorijo močan kompleks z molekulo 3.p. Medtem ko ohranjajo svoje osnovne spektralne lastnosti (glej Spektralna analiza), 3.p brez lipidov izgubijo številne funkcionalno pomembne, na primer sposobnost obnavljanja.
Čisti retinal ima rumeno barvo, maksimum njegovega absorpcijskega spektra je v območju 370 nm. Opsin je brezbarven, absorpcijski maksimum je v ultravijoličnem območju (cca. 280 nm). Barva molekule rodopsina je rdečkasto rožnata, maksimalni absorpcijski spekter je cca. 500 nm. Razlog za tako močan spektralni premik med nastajanjem kompleksa (od 370 do 500 nm – tako imenovani batokromni premik) še ni dobil nedvoumne razlage.
Maksimumi absorpcijskih spektrov rodopsinov in porfiropsinov pokrivajo precej široko območje vidnega spektra - od 433 do 562 nm za rodopsine in od 510 do 543 nm za porfiropsine. Če 3. čepke paglavca žabe, krapa in sladkovodne želve štejemo tudi za porfiropsine, tj. cianopsine z maksimalnim absorpcijskim spektrom pri 620 nm, potem se izkaže, da je to območje še širše. Razvoj mikrospektrofotometričnih metod je omogočil določitev absorpcijskih spektrov mnogih vrst posameznih fotoreceptorskih celic pri živalih in ljudeh. Glede na pridobljene podatke ima 3.p človeške mrežnice naslednje maksimume absorpcijskega spektra: palice 498, stožci, občutljivi na modro, zeleno in rdeče - 440, 535 in 575 nm.
V nemščini se je začel študij 3. str. raziskovalec H. Muller, ki je leta 1851 opisal, kako rožnato-vijolična mrežnica, izvlečena iz očesa žabe, na svetlobi postane rumenkasta in nato belkasta. Leta 1877 je F. Boll opisal tudi ta pojav in ugotovil, da je v vidnih celicah mrežnice nekakšna snov, občutljiva na rdečo svetlobo, in da je razbarvanje te snovi povezano z mehanizmom vida. Veliko zaslug za preučevanje 3. točk ima Kuhne (W. Kuhne, 1877), ki mu je uspelo 3. točke osamiti in jih podrobno preučiti. 3.p. je poimenoval vizualno vijolična, ugotovil njegovo proteinsko naravo, raziskal nekatere njegove spektralne lastnosti in fototransformacije ter odkril sposobnost 3.p., da se obnavlja v temi. K študiju 3. str. biokemik in fiziolog J. Wald.
Fototransformacije vidnih pigmentov. Ko je 3.p izpostavljen svetlobi, se v njih pojavi fotokemični cikel. transformacije, ki temeljijo na primarni fotokemični reakciji cis-trans izomerizacije mrežnice (glej Izomerija). V tem primeru je povezava med kromoforjem in proteinom motena. Zaporedje transformacij 3. str lahko predstavimo takole: rodopsin (kromofor je v cis obliki) -> prelumirodopsin -> lumirhodopsin -> metarodopsin I -> metarodopsin II -> protein opsin -> kromofor v trans obliki. Pod vplivom encima - retinol dehidrogenaze - se slednji pretvori v vitamin A, ki prihaja iz zunanjih segmentov palic in stožcev v celice pigmentne plasti mrežnice. Ko oko potemni, pride do regeneracije 3. p., za izvedbo reza je potrebna prisotnost cis-izomera vitamina A, ki služi kot začetni produkt za tvorbo kromofora (aldehid vitamina A). Če v telesu pride do pomanjkanja ali odsotnosti vitamina A, je lahko motena tvorba rodopsina in posledično se lahko razvije motnja vida v mraku, ti. nočna slepota (glej Hemeralopia). V procesu fototransformacij rodopsina na stopnji prehoda lumirhodopsina v metarodopsin I v receptorski celici se pojavi kot odgovor na svetlo bliskavico tako imenovanega. zgodnji (kratkolatenčni) receptorski potencial. Hkrati to ni vizualni signal, čeprav lahko služi kot eden od testov za preučevanje mehanizma transformacij 3.p v fotoreceptorski membrani. Funkcionalni pomen je t.i. pozni receptorski potencial, katerega latentna doba (5-10 ms) je primerljiva s časom nastanka metarodopsina II. Predpostavlja se, da reakcija prehoda metarodopsina I v metarodopsin II povzroči nastanek vizualni signal.
Ker se 3. p. na svetlobi nenehno obarvajo, morajo obstajati mehanizmi za njihovo stalno obnavljanje. Nekatere med njimi so izjemno hitre (fotoregeneracija), druge precej hitre (biokemične, regeneracijska, temna), tretje počasne (sinteza 3. p. med stalnim obnavljanjem fotoreceptorske membrane v vidni celici). Fotoregeneracija je fiziol, pomemben pri nevretenčarjih (npr. glavonožci- lignji, hobotnice). V mehanizmu biokem Pri regeneraciji 3. p pri vretenčarjih očitno igra pomembno vlogo encim izomeraza (glej), ki zagotavlja izomerizacijo trans-retinala (ali trans-vitamina A) ponovno v cis-izomerno obliko. Vendar dokončni dokazi o obstoju takega encima še niso na voljo. Sama reakcija tvorbe molekule 3.p v prisotnosti 11-cis-izomera retinala in opsina poteka enostavno, brez porabe energije. Odkrita je bila sposobnost beljenega rodopsina, da se podvrže reakciji fosforilacije (glej); domneva se, da je ta reakcija ena od povezav v mehanizmu svetlobne prilagoditve vidne celice.
Bibliografija: Akkerman Yu. Biofizika, trans. iz angleščine, M., 1964; Willie K. in Det e V. Biologija, prev. iz angleščine, M., 1974, bibliogr.; Konev S.V. in Volotovski I.D. Uvod v molekularno fotobiologijo, str. 61, Minsk, 1971; Ostrovsky M. A. in Fedorovich I. B. Foto-inducirane spremembe v fotoreceptorski membrani, v knjigi: Struktura in funkcije biol, membran, ed. A. S. Troshina et al., str. 224, M., 1975, bibliogr.; Fiziologija senzoričnih sistemov, ed. G. V. Gershuni, 1. del, str. 88, L., 1971; Biokemija in fiziologija vidnih pigmentov, ed. od H. Langer, V. a. o., 1973; Priročnik senzorične fiziologije, ed. od H. A. R. Jung a. o., v. 7, pt 1-2, B., 1972.
M. A. Ostrovskega.
Vizualni pigment strukturna in funkcionalna enota fotoobčutljive membrane fotoreceptorjev (Glej Fotoreceptorji) mrežnice - palice in stožci. V vidnem polju se pojavi prva stopnja vidnega zaznavanja - absorpcija kvantov vidne svetlobe. Molekulo Zp (molska masa približno 40.000) sestavljata kromofor, ki absorbira svetlobo, in opsin, kompleks beljakovin in fosfolipidov. Kromofor vseh mineralov je aldehid vitamina A 1 ali A 2 - retinal ali 3-dehidroretinal. Dve vrsti opsina (paličica in stožca) in dve vrsti retinala, če se združita v paru, tvorita 4 vrste opsina, ki se razlikujejo po absorpcijskem spektru: rodopsin (najpogostejši protein paličice) ali vizualno vijoličast (največja absorpcija 500 nm), jodopsin (562 nm), porfiropsin (522 nm) in cianopsin (620 nm). Primarna fotokemična povezava v mehanizmu vida (glej Vizija) je fotoizomerizacija mrežnice, ki pod vplivom svetlobe spremeni svojo ukrivljeno konfiguracijo v ravno. Tej reakciji sledi veriga temnih procesov, ki vodijo do pojava vizualnega receptorskega signala, ki se nato sinaptično prenese na naslednje živčne elemente mrežnice - bipolarne in horizontalne celice. Lit.: Fiziologija senzoričnih sistemov, 1. del, L., 1971, str. 88-125 (Priročnik za fiziologijo); Wald G., Molekularna osnova vizualnega vzbujanja, "Narava", 1968, v. 219. M. A. Ostrovskega.
Velika sovjetska enciklopedija. - M.: Sovjetska enciklopedija. 1969-1978 .
Oglejte si, kaj je "Vizualni pigment" v drugih slovarjih:
Strukturno in funkcionalno Enota je občutljiva na svetlobo. membrane retinalnih fotoreceptorjev paličic in stožcev. Molekula 3.p je sestavljena iz kromoforja, ki absorbira svetlobo, in kompleksa opsina iz beljakovin in fosfolipidov. Kromofor predstavlja aldehid vitamina A1... ... Biološki enciklopedični slovar
Rodopsin (vidno vijoličen) je glavni vidni pigment v paličicah mrežnice ljudi in živali. Nanaša se na kompleksne beljakovine - kromoproteine. Proteinske modifikacije, značilne za različne biološke vrste, se lahko zelo razlikujejo ... Wikipedia
VIZUALNI PIGMENT(I)- Glej fotopigment ... Slovar v psihologiji
Znotraj paličic je očesni pigment mrežnice, ki vključuje retinaldehid (retinal), vitamin A in beljakovine. Prisotnost rodopsina v mrežnici je nujna za normalen vid pri šibki svetlobi. Pod vplivom svetlobe ... ... Medicinski izrazi
RHODOPSIN, VIJOLIČNA VIZUALNA- (vizualno vijoličen) pigment mrežnice, ki ga vsebujejo palice, ki vključuje retinaldehid (retinal), vitamin A in beljakovine. Prisotnost rodopsina v mrežnici je nujna za normalen vid pri šibki svetlobi. Spodaj… … Razlagalni slovar medicine
- (vizualno vijolična), občutljiva na svetlobo. kompleksne beljakovine, osnovne vidni pigment paličastih celic mrežnice pri vretenčarjih in ljudeh. Z absorpcijo kvanta svetlobe (absorpcija največ pribl. 500 nm) R. razpade in povzroči vzbujanje... ... Naravoslovje. enciklopedični slovar
- (vidni pigment), fotoobčutljiva beljakovina paličic mrežnice vretenčarjev in vidnih celic nevretenčarjev. R. glikoprotein (mol. mas. Pribl. 40 tisoč; polipeptidna veriga je sestavljena iz 348 aminokislinskih ostankov), ki vsebuje... ... Kemijska enciklopedija
- (iz grške rhódon rose in opsis vid) vizualno vijolična, glavni vizualni pigment palic mrežnice vretenčarjev (razen nekaterih rib in dvoživk v zgodnjih fazah razvoja) in nevretenčarjev. Glede na kemično ... ... Velika sovjetska enciklopedija
- (vidno vijolična), fotoobčutljiva kompleksna beljakovina, glavni vidni pigment paličastih celic mrežnice pri vretenčarjih in ljudeh. Z absorpcijo kvanta svetlobe (absorpcija največ okoli 500 nm) rodopsin razpade in povzroči... ... enciklopedični slovar
Glavni članek: Palice (mrežnica) Rodopsin (zastarelo, a še vedno uporabljeno ime za vizualno vijolično) je glavni vidni pigment. Vsebujejo ga mrežnice morskih nevretenčarjev, rib, skoraj vseh kopenskih... ... Wikipedia
25-10-2014, 13:04
Svetlobne kvante absorbirajo v receptorjih specializirane molekule - vizualni fotopigmenti. Vidne pigmente sta neodvisno odkrila nemška fiziologa F. Boll in W. Kühne v letih 1877-1879. F. Boll je opazil, da je mrežnica, izolirana iz optične skodelice žabe, sprva videti rdeča, nato pa, ko zbledi na svetlobi, postane rumena in na koncu popolnoma brezbarvna.
W. Kuehne je ugotovil, da se pri normalno delujočem očesu živali barva mrežnice po intenzivni osvetlitvi s svetlobo obnovi, če žival ponovno postavimo v temo. Na podlagi teh opazovanj je bilo ugotovljeno, da je v mrežnici očesa svetlobno občutljiva snov (pigment), katere koncentracija se zmanjša na svetlobi in se obnovi v temi.
Vizualni pigmenti- to so kompleksne molekule kromolipoproteinov, ki jih pri vretenčarjih in nevretenčarjih sestavljata dva glavna dela: kromofor (barvni del molekule, ki določa barvo receptorja ob osvetlitvi) in beljakovina (opsin). Kromofor določa največjo in intenzivnost absorpcije svetlobe v pigmentu in je aldehid alkoholov - vitaminov A1 in A2.
Imena teh aldehidov sta retinal-1 in retinal-2. Prisotnost dodatne dvojne vezi v retinalu-2 vodi do premika maksimuma absorpcijskega spektra na daljše valovne dolžine. Mrežnica se veže na opsin, mutacijo proteina, ki jo najdemo samo v fotoreceptorjih. Opsin palice je skotopsin, opsin stožca pa fotopsin. Skotopsin je sestavljen iz 230-270 aminokislinskih ostankov, katerih zaporedje še ni določeno.
O fotopsinu je malo znanega: obstajajo le podatki o njegovi spiralni konfiguraciji. Dejanski del pigmentne molekule, ki absorbira svetlobo, retinal, je vrsta karotenoidnega pigmenta, ki je najpogostejši v fotosintezi. rastlinska in živalska tkiva.
Spektralne značilnosti pigmentov določajo kombinacije ene od mrežnic z eno ali drugo vrsto opsina, katerih raznolikost določa raznolikost vidnih pigmentov pri različnih živalih. Tako kombinacija skotopsina z retinalom-1 povzroči nastanek paličastega pigmenta - rodopsina z absorpcijskim maksimumom pri? = 500 nm, njegova kombinacija z retinalom-2 pa tvori še en paličasti pigment - porfiropsin z absorpcijskim maksimumom pri? =525 nm. Kombinacije retinal-1 z različni tipi fotopsini v mrežnici ljudi in opic tvorijo stožčaste pigmente z absorpcijskimi maksimumi pri? = 445 nm, ? = 535 nm in? =570 nm, imenovan cianolab, klorolab oziroma eritrolab.
Lastnosti pigmentov, ki absorbirajo svetlobo, so določene natančno z interakcijo iopsinskega kromoforja: absorpcijski maksimumi kromoforja in opsina, ločeno, so lokalizirani v regiji? =278 nm (retinal-1) in? = 378 nm (skotopsin), njihova spojina - rodopsin pa ima absorpcijski maksimum pri? = 500 nm. Največja absorpcija je eden od parametrov spektralnih karakteristik pigmenta, ki na splošno določa učinkovitost absorpcije svetlobnih kvantov (fotonov) s pigmentom. različne dolžine valovi (slika 3.1.4a).
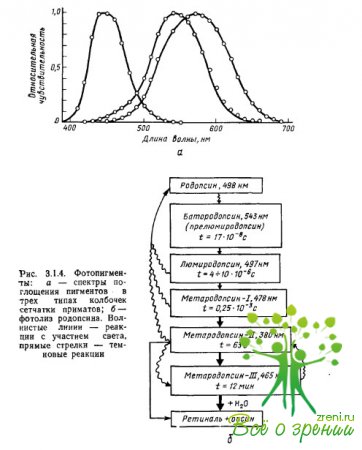
Prve meritve absorpcijskih spektrov pigmentov v posameznih stožcih opičje in človeške mrežnice so izvedli Brown, Wald in Marx et al., z uporabo mikrospektrofotometrične metode, posebej razvite za te namene. Preparat mrežnice smo postavili pod mikroskop, kar je omogočilo pod vizualnim nadzorom fokusiranje žarka svetlobe ene ali druge valovne dolžine na zunanji segment fotoreceptorja, kjer je pigment.
Nato so bile za vsako valovno dolžino ločeno izmerjene količine svetlobe, ki je prešla skozi stožec in je ta ni absorbiral (razlika med količino svetlobe, dovedene v stožec, in količino svetlobe, ki jo je prepustil, je značilna za učinkovitost pigmentna absorpcija fotonov dane valovne dolžine). Meritve smo ponovili, ko je pigment zbledel zaradi intenzivne osvetlitve (referenčne meritve). Kot rezultat odštevanja druge serije meritev od prve smo dobili diferencialni absorpcijski spekter pigmenta kot funkcijo svetlobne občutljivosti na valovno dolžino sevanja.
Iz sl. 3.1.4a je jasno, da imajo pigmenti širokopasovne, zelo prekrivajoče se absorpcijske spektre. Vsi fotopigmenti imajo dodatni absorpcijski maksimum, značilen za karotenoide (tako imenovani šis-vrh ali ?-vrh), ki je enak 2/3 glavnega maksimuma. Pri pigmentih, katerih glavni absorpcijski maksimumi so lokalizirani v kratko- in srednjevalovnih delih spektra, cis vrh pade v ultravijolično območje in ne vpliva na analizo sevanja iz vidnega spektra. Če nadalje pri konstruiranju funkcij spektralne občutljivosti pigmentov lestvico valovnih dolžin (os x) pretvorimo v frekvenčno lestvico, potem absorpcijski spektri vseh pigmentov, vključno z različnimi opsini, vendar enakega retiala (npr. retinal-1), bodo imele enako enako obliko.
S to predstavitvijo podatkov, ki jo je prvi predlagal Dartnall (1953) za pigmente na osnovi retinala-1, se absorpcijski spektri različnih fotopigmentov med seboj razlikujejo le v položaju maksimuma. In potem, če poznamo največjo absorpcijo pigmenta, lahko obnovimo obliko njegovega absorpcijskega spektra najprej v koordinatah "frekvenca sevanja - občutljivost", nato pa z uporabo razmerja c = ?f in v koordinatah "valovna dolžina - občutljivost".
Ta metoda rekonstrukcije absorpcijskega spektra neznanega pigmenta iz podatkov samo o njegovem maksimumu se imenuje metoda nomograma. Metoda nomograma se pogosto uporablja tudi v nevrofiziologiji barvnega vida pri odločanju o številu in vrsti fotodetektorjev, ki konvergirajo na določenem barvno občutljivem nevronu: spektralno občutljivost nevrona se primerja z enim ali drugim nomogramom (ali kombinacijo tega).
Raznolikost fotopigmentov v živalskem svetu je v korelaciji z raznovrstnostjo okoljskih razmer. Pri ribah in dvoživkah lahko mrežnica vsebuje tako rodopsin kot porfiropsin, katerih razmerje se tekom življenja posameznika spreminja; Pri anadromnih ribah je zamenjava rodopsina s porfiropsinom pred prehodom iz slane vode v sladko vodo in obratno. Povezava med naborom pigmentov in ekologijo habitata ni stroga: spremembe v sestavi pigmentov lahko povzročijo sezonske spremembe in cirkadiani ritmi.
Fotoizomerizacija.
Reakcija fotopigmenta na svetlobo je določena s številom absorbiranih kvantov, ne glede na valovno dolžino in energijo sevanja: na primer tistih, ki jih absorbira rodopsin. 10 kvantov svetlobe pri? =430 nm povzroči v njem enake strukturne spremembe (sl. 3.1.46) kot 10 absorbirane kvantne svetlobe pri? = 500 nm.
In spektralna selektivnost (absorpcijski spekter) pigmenta je določena le z različno verjetnostjo absorpcije svetlobnih kvantov iz različnih delov vidnega spektra. Tako vizualni pigmenti in navsezadnje fotoreceptorji delujejo kot »števci svetlobnih kvantov«, razlikujejo se le v verjetnosti absorbiranja fotonov različnih valovnih dolžin.
Ko ena molekula rodopsina absorbira 1 kvant svetlobe, pride do spremembe prostorske konfiguracije ene molekule mrežnice - fotoizomerizacije, ki je sestavljena iz transformacije 11-cm oblike mrežnice v popolnoma transformirano obliko (slika 3.1.4b).
Fotoizomerizacijo spremlja cikel fotokemičnih reakcij, za dešifriranje katerih je G. Wald leta 1930 prejel Nobelovo nagrado. Končna produkta fotoizomerizacije sta treksretinal in opsin. Vse vmesne reakcije do stopnje metarodopsina-P so reverzibilne: produkte teh reakcij je mogoče pretvoriti nazaj v rodopsin z reverzno gran-retinalno izomerizacijo.
To pomeni, da pred metarodopsinom-II v pigmentni molekuli ne pride do pomembnih konformacijskih sprememb. Na stopnji metarodopsina-II se retinal loči od proteinskega dela molekule. Od vseh stopenj izomerizacije le prva - pretvorba rodopsina v prelumirhodopsin (batorhodopsin) - zahteva svetlobo. Vse druge reakcije potekajo v temi. Fotoizomerizacija poteka podobno v stožčastih pigmentih.
Tako je fotoizomerizacija molekularni sprožilni mehanizem za fotorecepcijo, sestavljen iz spremembe prostorske konfiguracije molekule vidnega pigmenta. Slednje pojasnjuje, zakaj je narava izbrala karotenoide kot vidni pigment: prisotnost dvojnih vezi v retinalu določa možnost njegove izomerizacije in; njegova visoka reaktivnost. Okoli vsake dvojne vezi se lahko deli molekule vrtijo in upogibajo v prostoru. Ta oblika je najmanj stabilna in najbolj občutljiva na temperaturo in svetlobo. Najdeno v naravi različne vrste retinal - 7, 9, 11, 13-cis oblike, 9,13-dicis oblika, v vidnih pigmentih pa je retinal vedno v 11-cis obliki.
Resinteza pigmenta v fotoreceptorjih poteka neprekinjeno na svetlobi in v temi s sodelovanjem encima retinen izomeraze in je sestavljena iz povratne izomerizacije retinola iz transformacije v 11-cis obliko. V stožcih se ta encim nahaja v samih receptorjih, v paličicah, v pigmentnem epiteliju, kjer po ločitvi od opsina prehaja trans-retinol in kjer poteka njegova izomerizacija v 11-cis obliko. V tem primeru se retinal vitamina A ponovno uporabi, beljakovina pa se sintetizira v ribosomih notranjega segmenta.
Predpostavlja se, da pride do rekombinacije 11-cis-retinala z opsinom spontano. Približno 11 % vitamina A (retinola) je lokaliziranega v pigmentnem epiteliju, od 13 % v celotnem očesu. Vitamin A nastaja v jetrih tako, da verigo karotenoidov, ki jo najdemo v hrani, pretrgamo na dva dela in dodamo vodo.
Vitamin A se transportira s posebnim proteinom, ki veže retinol, do pigmentnega epitelija, kjer se oksidira v retinal. Pri pomanjkanju vitamina A se pojavi tako imenovana "nočna slepota" - zmanjšanje absolutne svetlobne občutljivosti, zlasti vpliva na vid v mraku, ki se spremeni v trajno slepoto zaradi uničenja beljakovine, ki je v prostem stanju nestabilna. Če imate nočno slepoto, je torej priporočljivo uživati korenje, ki vsebuje provitamin A – betakaroten.
Vsak fotoreceptor vsebuje samo en fotopigment, za katerega je značilen poseben absorpcijski spekter. V zvezi s tem ločimo kratko-, srednje- in dolgovalovne stožce, ki pri primatih vsebujejo pigmente z absorpcijskimi maksimumi pri 445 nm, 535 nm in 570 nm. U sladkovodne ribe Dolgovalovni stožec vsebuje pigment z absorpcijskim maksimumom pri 620 nm, kar je v korelaciji z obliko spektralne porazdelitve svetlobe v njihovem habitatu.
Včasih, po tradiciji, ki je ostala iz časa trikomponentne hipoteze Young-Helmholtza, se ti stožci imenujejo modro, zeleno in rdeče občutljivi receptorji. Toda od druge polovice 20. stoletja, ko je bilo ugotovljeno, da noben fotoreceptor sam ne izvaja spektralne analize, ampak reagira le na število absorbiranih fotonov, ne glede na valovno dolžino sevanja, se ta barvna imena receptorjev uporabljajo le v metaforičen smisel.
Razvrstitev palic, na primer pri žabi, na "zelene" in "rdeče" v nekaterih primerih ne temelji na spektralni občutljivosti pigmenta, ki ga vsebujejo, temveč na barvi receptorskih teles v prepuščeni svetlobi. Tako "zelene" palice vsebujejo pigment z največjo absorpcijo pri 432 nm, zato ob osvetlitvi z belo svetlobo absorbirajo kratkovalovno sevanje, prenašajo vse druge valovne dolžine, pri vizualnem pregledu pa so videti zelenkaste.
V mrežnici rib je bila opažena povezava med dolžino stožca in spektralno vrsto pigmenta, ki ga vsebuje: najkrajši med njimi vsebuje kratkovalovni pigment, najdaljši pa dolgovalovni. Eberly je predlagal, da je to morda eden od mehanizmov za boj proti kromatski aberaciji. Zaradi kromatske aberacije se ostra slika srednjevalovnega sevanja nahaja globlje (dlje od leče) kot ostra slika kratkovalovnega sevanja, žarišče za dolgovalovno sevanje pa je lokalizirano še dlje.
Dovolj je, da kratko-, srednje- in dolgovalovne receptorje postavimo na ustrezne nivoje, tako da so vsi deli spektralnega sevanja usmerjeni na receptorje enako ostro. To lahko pojasni dejstvo, da v mrežnici rib enojni stožci s kratkimi segmenti vsebujejo kratkovalovni pigment, v dvojnem stožcu pa najdaljši segment vsebuje dolgovalovni pigment, krajši pa kratko- ali srednjevalovni pigment. . Dvojni stožci z enako dolgimi segmenti vsebujejo enak pigment.
ωπς - oko) - pogosto ime več vidnih pigmentov ljudi in nekaterih sesalcev. Ti pigmenti so sestavljeni iz beljakovinske molekule, vezane na kromoforno mrežnico. Vsebuje se v stožcih mrežnice in zagotavlja barvni vid, v nasprotju z drugim vidnim pigmentom - rodopsinom, ki je odgovoren za vid v somraku.Ugotovljeno je bilo, da vizualni pigmenti vključujejo opsine. Različni opsini se razlikujejo po aminokislinah v svojih molekulah in absorbirajo svetlobo pri nekoliko različnih valovnih dolžinah, kot molekule, vezane na mrežnico.
Obstoj stožčastega pigmenta je prvi (posredno) odkril George Wald, ki mu je dal ime jodopsin. . Za ta dela je leta 1967 prejel Nobelovo nagrado za fiziologijo in medicino.
Helmholtz je tudi verjel, da bi morale biti v "končnem aparatu optičnih živčnih vlaken" (fotoreceptorji mrežnice) tri vrste fotokemično razgradljivih snovi, ki imajo različno občutljivost na različne dele spekter .
Po trikomponentni teoriji vida, ki prevladuje v sodobni znanosti, se domneva, da obstajajo tri vrste tega pigmenta in da mrežnica vsebuje tri vrste stožcev, ki so občutljivi na modro, zeleno in rdečo svetlobo. V skladu s tem je jodopsin, ki se nahaja v stožcih očesa, sestavljen iz treh pigmentov - klorolaba, eritrolaba in cianolaba; prvi od njih absorbira žarke, ki ustrezajo rumeno-zelenemu (absorpcijski pas 450-630 nm), drugi - rumeno-rdeči (500-700 nm) in tretji - modro-zeleni (500-700 nm) del vidnega dela svetlobe. spekter.
Vrste vizualnih pigmentov
Prve poskuse odkritja treh pigmentov in, kot je bilo pričakovano, treh vrst stožcev (na podlagi predpostavke hipoteze o trikomponentnem vidu, da vsak stožec vsebuje samo en pigment) je izvedel Rushton, ki je izpopolnil tehniko denzitometrije za intravitalno merjenje. absorpcijskih koeficientov svetlobe različnih valovnih dolžin - v plasti mrežničnih fotoreceptorjev. Dokazano je, da pri barvnih anomalijah manjka eden od pigmentov, ki jih najdemo pri ljudeh z normalnim vidom: "eritrolab" (največ približno 585 nm) pri protanopu in "klorolab" (največ približno 540 nm) pri deuteranopu.
Upoštevati je treba, da se pri uporabi izraza "sprejemnik" v nekaterih primerih celoten niz fotoreceptorjev z enako spektralno občutljivostjo šteje za enega. sprejemnik; v drugih primerih pa vprašanje, ali vsak fovealni stožec vsebuje tri sprejemnik ali samo enega. Hkrati ni kršena strogost pojma "sprejemnik", ki v tem primeru ne vključuje posebnih morfoloških značilnosti.
Naslednji korak v tej smeri je bila študija fotopigmentov v posameznih stožcih ljudi in makakov. Fovealni stožci so premajhni, da bi jih preučevali, vsi pridobljeni podatki pa se nanašajo samo na parafovealne stožce. Vsak stožec, vsaj ekstrafovealni, očitno vsebuje samo enega od pigmentov ali pretežno enega od njih.
Sodobne metode preučevanja stožčastih fotopigmentov
Denzitometrija celic mrežnice
→ denzitometrija, je lahko naslednja:
Če je prve poskuse najti tri pigmente in, kot je bilo pričakovano, tri vrste stožcev izvedel Rushton, ki je tehniko izpopolnil Denzitometrija za merjenje absorpcijskih koeficientov svetlobe z različnimi valovnimi dolžinami - v plasti fotoreceptorjev mrežnice, in pokazalo se je, da pri barvnih anomalijah manjka eden od pigmentov, ki jih najdemo pri ljudeh z normalnim vidom: "eritrolab" (največ približno 585 nm.) v protanope in "klorolab" (največ približno 540 nm.) - v deuteranopu lahko zdaj s pomočjo posebnih dinestometričnih metod z uporabo sodobnih denzitometrov znanstveniki določijo delovanje stožcev in palic v stanju normalne aktivnosti in diagnosticirajo njihove bolezni.
Poglej tudi
Viri
Literatura
- Khokhlova T.V. Sodobne ideje o viziji sesalcev // Journal of General Biology. Letnik 73, 2012. št. 6, november-december. Stran 418-434.


