vizualni pigmenti. Rudimentarni del povrhnjice kože je sestavljen iz plasti. Dnevni in nočni vid
Vidni pigmenti so koncentrirani v membranah zunanjih segmentov. Vsaka palica vsebuje približno 10 8 pigmentnih molekul. Organizirani so v več sto ločenih diskov (približno 750 v opičji palici), ki niso povezani z zunanjo membrano. V stožcih se pigment nahaja v posebnih pigmentnih gubah, ki so nadaljevanje zunanje celične membrane fotoreceptorja. Pigmentne molekule sestavljajo približno 80 % vseh beljakovin diska. Vidni pigmenti so tako gosto zapakirani v membranah zunanjega segmenta, da razdalja med dvema molekulama vidnega pigmenta v palici ne presega 10 nm. Tako gosto pakiranje poveča verjetnost, da bo foton svetlobe, ki gre skozi plast fotoreceptorskih celic, zajet. Postavlja se naslednje vprašanje: kako nastanejo signali, ko svetlobo absorbirajo vidni pigmenti?
Absorpcija svetlobe z vidnimi pigmentiDogodke, ki se zgodijo, ko svetlobo absorbira palični pigment rodopsin, so preučevali s psihofiziološkimi, biokemičnimi in molekularnimi tehnikami. Molekula vidnega pigmenta je sestavljena iz dveh komponent: proteina, imenovanega opsin, in kromofora, 11-cis-vitamin A-aldehida, imenovanega retinal (slika 1). Treba je pojasniti, da kromofor vsebuje kemično skupino, ki daje spojini barvo. Kvantitativne značilnosti absorpcijske sposobnosti pigmentov smo proučevali s spektrofotometrijo. Ko smo rodopsin, vidni pigment paličic, osvetlili s svetlobo različnih valovnih dolžin, se je najbolje absorbirala modro-zelena svetloba z valovno dolžino okoli 500 nm. Podoben rezultat je bil dosežen tudi z osvetlitvijo ene paličice pod mikroskopom s svetlobnimi žarki različnih valovnih dolžin. Je bil razkrit zanimiva odvisnost med absorpcijskim spektrom rodopsina in našim zaznavanjem somračne svetlobe. Kvantitativne psihofizične študije, opravljene na ljudeh, so pokazale, da je modrikasto zelena dnevna svetloba z valovno dolžino okoli 500 nm optimalna za zaznavo svetlobe somraka v temi. Čez dan, ko so paličice neaktivne in se uporabljajo le stožci, smo najbolj občutljivi na rdečo barvo, ki ustreza absorpcijskemu spektru stožcev (o tem bomo govorili kasneje).
Ko en foton absorbira rodopsin, se retinal fotoizomerizira in preide iz 11-cis v trans konfiguracijo. Ta prehod se zgodi zelo hitro: v približno 10-12 sekundah. Po tem je tudi beljakovinski del pigmenta podvržen vrsti transformacijskih sprememb, pri čemer nastanejo številni vmesni produkti. Ena od konformacij proteinskega dela, metarodopsin II, je najpomembnejša za prenos signala (o tem bomo razpravljali pozneje v tem poglavju). Slika 2 prikazuje zaporedje dogodkov med razbarvanjem in regeneracijo aktivnega rodopsina. Metarhodopsin II nastane po 1 ms. Regeneracija pigmenta po njegovem razpadu poteka počasi, v nekaj minutah; to zahteva transport mrežnice od fotoreceptorjev do pigmentnega epitelija.
Struktura rodopsinaNa molekularni ravni je opsin protein sestavljen iz 348 aminokislinskih ostankov, ki tvorijo 7 hidrofobnih con, od katerih je vsaka sestavljena iz 20-25 aminokislin, ki sestavljajo 7 transmembranskih vijačnic. N-konec molekule se nahaja v zunajceličnem prostoru (tj. znotraj diska paličice), C-konec pa se nahaja v citoplazmi.
V temi je 11-cis-retinal tesno vezan na protein opsin. Zajetje fotona vodi do izomerizacije vsega cis retinala v retinalnem tronu. V tem primeru se kompleks opsin all-tron-retinal hitro spremeni v metarodopsin II, ki disociira na opsin in all-tron retinal. Regeneracija rodopsina je odvisna od interakcije fotoreceptorjev in pigmentnih celic. Metarhodopsin II aktivira in vzdržuje drugi messenger sistem.
Retinal je povezan z opsinom preko ostanka lizina, ki se nahaja v sedmem transmembranskem segmentu. Opsin spada v družino proteinov s 7 transmembranskimi domenami, ki vključuje tudi metabotropne mediatorske receptorje, kot so adrenergični in muskarinski receptorji. Tako kot rodopsin tudi ti receptorji signalizirajo sekundarnim posrednikom prek aktivacije G-proteina. Rhodopsin je izjemno stabilen v temi. Bayor je izračunal, da spontana toplotna izomerizacija molekule rodopsina traja približno 3000 let ali 10 23 več kot za fotoizomerizacijo.
vizualni pigmenti
vizualni pigmenti
Struktura rodopsina
Stožci in barvni vid
barvna slepota
Literatura
vizualni pigmenti
Vidni pigmenti so koncentrirani v membranah zunanjih segmentov. Vsaka palica vsebuje približno 10 8 pigmentnih molekul. Organizirani so v več sto ločenih diskov (približno 750 v opičji palici), ki niso povezani z zunanjo membrano. V stožcih se pigment nahaja v posebnih pigmentnih gubah, ki so nadaljevanje zunanje celične membrane fotoreceptorja. Pigmentne molekule sestavljajo približno 80 % vseh beljakovin diska. Vidni pigmenti so tako gosto zapakirani v membranah zunanjega segmenta, da razdalja med dvema molekulama vidnega pigmenta v palici ne presega 10 nm. Tako gosto pakiranje poveča verjetnost, da bo foton svetlobe, ki gre skozi plast fotoreceptorskih celic, zajet. Postavlja se naslednje vprašanje: kako nastanejo signali, ko svetlobo absorbirajo vidni pigmenti?
Absorpcija svetlobe z vidnimi pigmenti
Dogodke, ki se zgodijo, ko svetlobo absorbira palični pigment rodopsin, so preučevali s psihofiziološkimi, biokemičnimi in molekularnimi tehnikami. Molekula vidnega pigmenta je sestavljena iz dveh komponent: proteina, imenovanega opsin, in kromofora, 11-cis-vitamin A-aldehida, imenovanega retinal (slika 1). Treba je pojasniti, da kromofor vsebuje kemično skupino, ki daje spojini barvo. Kvantitativne značilnosti absorpcijske sposobnosti pigmentov smo proučevali s spektrofotometrijo. Ko smo rodopsin, vidni pigment paličic, osvetlili s svetlobo različnih valovnih dolžin, se je najbolje absorbirala modro-zelena svetloba z valovno dolžino okoli 500 nm. Podoben rezultat je bil dosežen tudi z osvetlitvijo ene paličice pod mikroskopom s svetlobnimi žarki različnih valovnih dolžin. Ugotovljeno je bilo zanimivo razmerje med absorpcijskim spektrom rodopsina in našim zaznavanjem somračne svetlobe. Kvantitativne psihofizične študije, opravljene na ljudeh, so pokazale, da je modrikasto zelena dnevna svetloba z valovno dolžino okoli 500 nm optimalna za zaznavo svetlobe somraka v temi. Čez dan, ko so paličice neaktivne in se uporabljajo le stožci, smo najbolj občutljivi na rdečo barvo, ki ustreza absorpcijskemu spektru stožcev (o tem bomo govorili kasneje).
Ko en foton absorbira rodopsin, se retinal fotoizomerizira in preide iz 11-cis v trans konfiguracijo. Ta prehod se zgodi zelo hitro: v približno 10-12 sekundah. Po tem je tudi beljakovinski del pigmenta podvržen vrsti transformacijskih sprememb, pri čemer nastanejo številni vmesni produkti. Ena od konformacij proteinskega dela, metarodopsin II, je najpomembnejša za prenos signala (o tem bomo razpravljali pozneje v tem poglavju). Slika 2 prikazuje zaporedje dogodkov med razbarvanjem in regeneracijo aktivnega rodopsina. Metarhodopsin II nastane po 1 ms. Regeneracija pigmenta po njegovem razpadu poteka počasi, v nekaj minutah; to zahteva transport mrežnice od fotoreceptorjev do pigmentnega epitelija.
Struktura rodopsina
Na molekularni ravni je opsin protein sestavljen iz 348 aminokislinskih ostankov, ki tvorijo 7 hidrofobnih con, od katerih je vsaka sestavljena iz 20-25 aminokislin, ki sestavljajo 7 transmembranskih vijačnic. N-konec molekule se nahaja v zunajceličnem prostoru (tj. znotraj diska paličice), C-konec pa se nahaja v citoplazmi.
Slika 1. Struktura vretenčarskega rodopsina, vgrajenega v fotoreceptorsko membrano. Spirala je nekoliko odvita, da pokaže lokacijo mrežnice (označeno s črno). C - C-konec, N - N-konec.
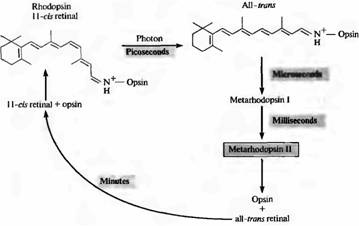
Slika 2. Cvetenje rodopsina na svetlobi. V temi je 11-cis-retinal tesno vezan na protein opsin. Zajetje fotona vodi do izomerizacije vsega cis retinala v retinalnem tronu. V tem primeru se kompleks opsin all-tron-retinal hitro spremeni v metarodopsin II, ki disociira na opsin in all-tron retinal. Regeneracija rodopsina je odvisna od interakcije fotoreceptorjev in pigmentnih celic. Metarhodopsin II aktivira in vzdržuje drugi messenger sistem.
Retinal je povezan z opsinom preko ostanka lizina, ki se nahaja v sedmem transmembranskem segmentu. Opsin spada v družino proteinov s 7 transmembranskimi domenami, ki vključuje tudi metabotropne mediatorske receptorje, kot so adrenergični in muskarinski receptorji. Tako kot rodopsin tudi ti receptorji signalizirajo sekundarnim posrednikom prek aktivacije G-proteina. Rhodopsin je izjemno stabilen v temi. Bayor je izračunal, da spontana toplotna izomerizacija molekule rodopsina traja približno 3000 let ali 10 23 več kot za fotoizomerizacijo.
Stožci in barvni vid
Osupljive raziskave in poskusi, ki sta jih v 19. stoletju izvajala Young in Helmholtz, so opozorili na zelo pomembno vprašanje o barvnem vidu, znanstveniki pa so sami dali jasno in natančno razlago tega pojava. Njihov sklep o obstoju treh različne vrste barvnih fotoreceptorjev je prestala preizkus časa in je bila nato potrjena na molekularni ravni. Spet lahko citiramo Helmholtza, ki je primerjal zaznavo svetlobe in zvoka, barve in zvočnega tona. Lahko mu zavidamo jasnost, moč in lepoto njegove misli, zlasti v primerjavi z zmedenimi vitalističnimi koncepti, ki so bili razširjeni v 19. stoletju:
Vse razlike v barvnih tonih so odvisne od kombinacije v različnih razmerjih treh osnovnih barv ... rdeče, zelene in vijolične ... Tako kot je zaznavanje sončne svetlobe in njene toplote odvisno ... od tega, ali sončni žarki zadenejo živcev, ki prihajajo iz receptorjev za vid ali iz receptorjev za toplotno občutljivost. Kot je predlagal Young v svoji hipotezi, je razlika v zaznavanju različnih barv odvisna preprosto od tega, kateri od treh vrst fotoreceptorjev je bolj aktiviran s to svetlobo. Ko so vse tri vrste enako navdušen postane bel ...
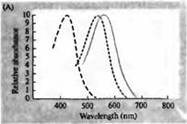
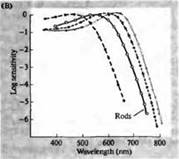
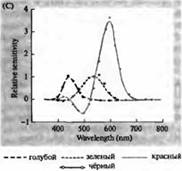
riž. 3. Spektri občutljivosti človeških fotoreceptorjev in razni vizualni pigmenti. (A) Krivulje občutljivosti treh barvnih vizualnih pigmentov, ki prikazujejo vrhove absorpcije pri valovnih dolžinah, ki ustrezajo cianu, zeleni in rdeči. (B) Modri, zeleni in rdeči spektri občutljivosti stožcev in palic (prikazano črno) pri makakih. Odzivi so bili zabeleženi s sesalnimi elektrodami, povprečeni in normalizirani. Krivulje paličastega spektra so bile pridobljene pri študiju vidnih pigmentov pri ljudeh. (C) Primerjava opičjega in človeškega stožčastega spektra z uporabo testa barvne občutljivosti. Neprekinjena krivulja prikazuje poskus za določanje občutljivosti na barve pri ljudeh, ko so izpostavljeni svetlobi različnih valovnih dolžin. Črtkana črta prikazuje rezultate, predvidene na podlagi registracije tokov v posameznih stožcih, po korekciji za absorpcijo svetlobe v leči in pigmentih na poti do zunanjega segmenta. Ujemanje rezultatov obeh poskusov je presenetljivo visoko.
Če na bel zaslon hkrati projiciramo dva svetlobna snopa različnih barv ... vidimo samo eno barvo, bolj ali manj drugačno od obeh teh barv. Izjemno dejstvo, da lahko z mešanico treh osnovnih barv zaznamo vse odtenke v sestavi zunanje svetlobe, lahko bolje razumemo, če oko primerjamo s suhim ... Pri zvoku ... slišite daljše valove kot nizke tone in kratke valove - kot visoke in prodorne, poleg tega je uho sposobno zajeti veliko zvočnih valov hkrati, tj. veliko opomb. Vendar pa se β v tem primeru ne zlijejo v en kompleksen akord, tako kot se različne barve ... zlijejo v eno kompleksno barvo. Oko ne more opaziti razlike, če zamenjamo oranžna barva do rdeče ali rumene barve; če pa slišimo noti do in mi zveneti hkrati, se nam tak zvok ne zdi kot nota re. Če bi uho zaznavalo glasbene tone tako, kot oko zaznava barve, bi lahko vsak akord predstavili s kombinacijo treh konstantnih not, ene zelo nizke, ene zelo visoke in ene vmesne, pri čemer bi ustvarili vse možne glasbene učinke le s spreminjanjem relativne glasnosti teh. tri note ... Vendar pa lahko vidimo gladek prehod barv iz ene v drugo skozi neskončno število odtenkov in gradacij ... Način, kako zaznavamo vsako barvo ... je odvisen predvsem od strukture našega živčni sistem. Treba je priznati, da trenutno niti pri ljudeh niti pri tetrapodih ni opisana anatomska osnova, ki bi potrdila teorijo zaznavanja barv.
Te natančne in daljnovidne napovedi so bile potrjene z vrsto različnih opazovanj. Z uporabo spektrofotometrije so Wald, Brouck, McNicol in Dartnell et al pokazali prisotnost treh vrst stožcev z različnimi pigmenti v človeški mrežnici. Baylorju in kolegom je tudi uspelo preusmeriti tokove iz stožcev opic in ljudi. Ugotovljeno je bilo, da imajo tri populacije stožcev različne, a prekrivajoče se razpone občutljivosti na modri, zeleni in rdeči del spektra. Optimalne valovne dolžine za vzbujanje električnih signalov so natančno sovpadale z vrhovi absorpcije svetlobe z vidnimi pigmenti, ugotovljenimi s spektrofotometrijo in psihofizičnimi eksperimenti za merjenje občutljivosti očesa na barvni spekter. Končno je Natais kloniral in sekvenciral gene, ki kodirajo pigment opsin v treh vrstah stožcev, ki so občutljivi na rdečo, zeleno in modro.
Kako so torej molekule različnih vidnih pigmentov sposobne prednostno ujeti svetlobo določene valovne dolžine? Izkazalo se je, da je rodopsin vidni pigment paličic in vsi trije vidni pigmenti stožcev vsebujejo isti kromofor, 11-cis-retinal. Vendar se aminokislinska zaporedja beljakovinskega dela pigmenta med seboj razlikujejo. Razlike v samo nekaj aminokislinah pojasnjujejo njihovo različno občutljivost na spekter.
barvna slepota
Čeprav ena vrsta fotoreceptorjev ni sposobna zaznati barve sama, tri vrste stožcev, kot je prikazano na sl. 4, že zmožen.
Za razpoznavo barv bi načeloma zadoščali dve vrsti stožcev z različnimi pigmenti, vendar bi v tem primeru več kombinacij valovnih dolžin zaznali enako. Podobna situacija se pojavi, ko oseba trpi za barvno slepoto. Takšni ljudje, kot je pokazal Nathans, imajo genetske okvare, ki vodijo v odsotnost enega od pigmentov. Z vidika sedanjega stanja znanosti ne moremo kaj, da ne bi bili presenečeni nad tem, kako lepo molekularni mehanizmi potrjujejo briljantno in presenetljivo natančno razmišljanje Younga in Helmholtza.
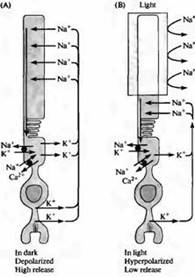
riž. 4. "Temni" tok v palici. (A) V temi gredo natrijevi ioni skozi kationske kanale v zunanjem segmentu paličic, kar povzroči depolarizacijo; skozi te kationske kanale lahko prehajajo tudi kalcijevi ioni. Tokovna zanka poteka skozi ožino palice zaradi dejstva, da kalijev tok teče navzven v notranjem segmentu membrane. (B) Ko je zunanji segment osvetljen, se kanalčki zaprejo zaradi zmanjšanja znotrajcelične koncentracije cGMP in paličica se hiperpolarizira. Hiperpolarizacija povzroči zmanjšanje sproščanja nevrotransmiterja. Koncentracijo natrija, kalija in kalcija v paličici vzdržujejo posebne črpalke in ionski izmenjevalci, ki se nahajajo v območju notranjega segmenta (črni krogi). V zunanjem segmentu najdemo tudi prenašalce kalcija.
Njihovo zamisel, da naj bi se glavni atributi barvnega vida in barvne slepote nahajali v samih fotoreceptorjih, so potrdili z neposrednimi fiziološkimi meritvami, pa tudi s proučevanjem razlik v strukturi pigmentov na genetski in beljakovinski ravni, potencial je reda velikosti. -40 mV, kar je daleč od ravnotežnega kalijevega potenciala E K, ki je - 80 mV.

riž. 5. Vloga cGMP pri odpiranju natrijevih kanalčkov v zunanjem segmentu paličic. Aktivnost posameznih kanalov je bila zabeležena s pomočjo objemke navznoter, ki je bila v raztopini z različnimi koncentracijami cGMP. Odpiranje kanala povzroči odklon toka navzgor. Frekvenca odpiranja kanalov je bila v kontroli zelo nizka, dodatek cGMP je povzročil povečanje frekvence odpiranja posameznega kanala, ki je bila neposredno odvisna od koncentracije cGMP.
Vhodni tok v temi prenašajo predvsem natrijevi ioni po elektrokemičnem gradientu skozi kationske kanale zunanjega segmenta. Hiperpolarizacija fotoreceptorja pod vplivom svetlobe je posledica zaprtja teh kanalov, kar vodi do premika membranskega potenciala proti E K.
Lastnosti fotoreceptorskih kanalov
Kationski kanali zunanjega segmenta imajo v fizioloških pogojih razmerje v prevodnosti kalcijevih / natrijevih / kalijevih ionov 12,5: 1,0: 0,7, prevodnost posameznega kanala pa je približno 0,1 pS 21). Ker je koncentracija natrija veliko višja od koncentracije kalcija, približno 85 % vhodnega toka predstavljajo natrijevi ioni. Gonilna sila kalijevih ionov je usmerjena izven celice. Ko kalcijevi ioni prehajajo skozi kanal, so ti ioni trdno vezani na steno pore in tako motijo prevajanje drugih ionov. Zaradi tega odstranitev kalcija iz zunajceličnega medija povzroči lažji prehod kalijevih in natrijevih ionov skozi kanale, katerih prevodnost se poveča na 25 pS.
Fesenko, Yau, Baylor, Strayer in sodelavci so pokazali, da ciklični HMP igra vlogo notranjega nosilca signala od diska do površine membrane. Kot je prikazano na sliki 4, visoka koncentracija cGMP v citoplazmi ohranja kationske kanale odprte. Z zmanjšanjem koncentracije cGMP z notranje površine membrane postane odprtje kationskih kanalov redek dogodek. Tako je membranski potencial fotoreceptorjev odraz koncentracije cGMP v citoplazmi: večja ko je koncentracija cGMP, večja je depolarizacija celice. Koncentracija cGMP pa je odvisna od intenzivnosti vpadne svetlobe. Povečanje jakosti svetlobe povzroči zmanjšanje koncentracije cGMP in zmanjša delež odprtih kanalov. V odsotnosti cGMP so skoraj vsi kanali zaprti, odpornost membrane zunanjega segmenta pa se približa odpornosti lipidnega dvosloja.
Molekularna struktura cGMP-odvisnih kanalčkov
Za ionske kanale zunanjega segmenta paličic smo izolirali cDNA in določili aminokislinska zaporedja podenot teh kanalov v človeški, goveji, mišji in piščančji mrežnici. Ugotovljena je bila pomembna podobnost DNK za te in druge ionske kanale, odvisne od cGMP – kot so tisti, ki jih najdemo v vohalnih
Transdukcija signala v fotoreceptorjih
Kako fotoizomerizacija rodopsina povzroči spremembo membranskega potenciala? Z leti je bilo ugotovljeno, da je za vzbujanje električnih signalov v paličicah in stožcih potreben nekakšen notranji posrednik. Eden od razlogov za sum, da informacije o absorpciji fotonov v območju zunanjega segmenta paličice prenaša nosilec, je bilo dejstvo, da se sam pigment rodopsin nahaja znotraj diska, signal pa se širi skozi citoplazmo do zunanjo membrano. Drugi razlog je bil precejšen porast odziva. Baylor in sodelavci, ki so preučevali fotoreceptorje pri želvah, so pokazali, da do zmanjšanja prevodnosti membrane in zabeleženih električnih signalov pride že, ko se absorbira samo en foton in se aktivira ena od 10 8 pigmentnih molekul.
Zaporedje dogodkov, v katerih aktivirana molekula fotopigmenta spremeni membranski potencial, je bilo razjasnjeno s preučevanjem zunanjih segmentov paličic in stožcev z uporabo dvoelektrodnega vpenjanja potenciala in tehnik molekularne biologije. Shema prenosa signala od absorpcije fotona svetlobe do električnega signala je prikazana na sl. 3.
V temi v zunanjih segmentih paličic in stožcev teče stalen vhodni "temni" tok. Rezultat je njihov membranski sistem. Membranske regije teh kanalov kažejo strukturne podobnosti z drugimi kationsko selektivnimi kanali, zlasti v regiji S4 in v regiji, ki tvori ionsko poro. Fotoreceptorski ionski kanali so tetrameri, sestavljeni iz vsaj 2 različnih beljakovinskih podenot z molekulsko maso 63 oziroma 240 kDa.
Znotrajcelična vezavna mesta za nukleotide se nahajajo blizu karboksilnega konca podenot. Izražanje teh podenot v jajčnih celicah vodi do tvorbe kationskih kanalov z lastnostmi, podobnimi tistim, ki se nahajajo v zunanjih segmentih paličic: aktivira jih cGMP in imajo pričakovana razmerja prevodnosti in prepustnosti.

riž. 6. Mehanizem aktivacije G-proteina ob aktivaciji molekule fotopigmenta. G-protein transducin veže GTP v prisotnosti metarodopsina II, kar vodi do aktivacije fosfodiesteraze, ta pa hidrolizira cGMP. Ko koncentracija cGMP pade, se natrijevi kanalčki zaprejo.
Presnovna kaskada cikličnega GMP
Veriga dogodkov, ki vodijo do zmanjšanja koncentracije cGMP in posledično zaprtja ionskih kanalov, je prikazana na sliki 5. Zmanjšanje znotrajcelične koncentracije cGMP povzroči svetloba, kar povzroči nastanek metarhodopsina II, vmesnega produkta pri razpadu fotopigmenta. Metarhodopsin II deluje na transducin G-proteina, ki je sestavljen iz 3 polipeptidnih verig.
Medsebojno delovanje metarhodopsina II in transducina vodi do zamenjave molekule GDP, vezane na G-protein, z GTP. To aktivira podenoto G-proteina, ki se loči od podenot in posledično aktivira fosfodiesterazo, ki se nahaja v membranski regiji: encim, ki hidrolizira cGMP. Koncentracija cGMP pade, odprtih ionskih kanalov je manj, paličica postane hiperpolarizirana. Kaskado prekine fosforilacija C-konca aktivnega metarodopsina II. Ključno vlogo cGMP pri uravnavanju stanja kationskih kanalov so potrdili biokemični poskusi. Osvetlitev fotoreceptorjev lahko povzroči 20% padec ravni cGMP v celici.
Receptorji vretenčarjev, ki se depolarizirajo ob izpostavljenosti svetlobi
Zanimiva izjema zgoraj navedenega mehanizma fotorecepcije so nekateri receptorji vretenčarjev. Kuščarji imajo tretje oko, ki se nahaja na vrhu glave. Vsebuje majhne "stožce", ki so sposobni zaznati sliko, podobno tisti, ki jo zaznavajo glavne (stranske) oči. Ti fotoreceptorji pa so izjemni, saj se ob osvetlitvi depolarizirajo. Kanali, odvisni od nukleotidov, imajo tukaj podobno strukturo in funkcijo kot fotoreceptorji drugih vretenčarjev, z eno izjemo: aktivacija fotoreceptorja in G-proteina vodi do povečanja koncentracije cGMP. Posledično se odprejo kanali zunanjega segmenta in kationi se usmerijo v celico in tvorijo "svetlobni" tok. To je posledica zaviranja aktivnosti fosfodiesteraze v temi. Posledično se v očesu kuščarja pojavi naslednje zaporedje dogodkov: svetloba - [cGMP] - odpiranje kationskih kanalov zunanjega segmenta - depolarizacija.
Ojačitev signala v kaskadi cGMP
Dvostopenjska kaskada cGMP zagotavlja znatno ojačanje prvotnega signala, kar pojasnjuje izjemno občutljivost paličic na svetlobo. Prvič, ena molekula aktivnega metarodopsina II katalizira dodajanje številnih molekul GTP namesto GDP in tako sprosti na stotine podenot G-proteina. Drugič, vsaka podenota aktivira eno molekulo fosfodiesteraze v disku, ki je sposobna odcepiti ogromno število molekul cGMP, ki se nahajajo v citoplazmi, in tako povzročijo zaprtje velikega števila kanalov.
Signali kot odziv na posamezne kvante svetlobe
Podatki, da lahko posamezni svetlobni kvanti povzročijo zaznaven občutek svetlobe, ki jo povzroči veliko število vprašanja. Kako velik je ta en sam odgovor? Kako ta signal izstopa od ravni hrupa? In kako se te informacije zanesljivo prenašajo iz mrežnice v višje vidne centre? Da bi izmerili signale kot odziv na posamezne kvante svetlobe, so Baylor in sodelavci posneli tokove iz posameznih palic v mrežnici krastač, opic in ljudi. Ti poskusi so edinstven primer eksperimenta, ki prikazuje, kako tak težak proces, kot zaznavanje šibkih svetlobnih utrinkov, lahko korelira s spremembami, ki se dogajajo na ravni posameznih molekul.
Postopek za izolacijo fragmenta mrežnice iz živalskega ali trupelnega materiala je treba za te poskuse izvesti v temi. Za merjenje toka se zunanji segment palice posesa v tanko pipeto. Kot je bilo pričakovano, ti poskusi kažejo, da v temi tok nenehno teče znotraj zunanjega segmenta. Utripi svetlobe zaprejo kanale v zunanjem segmentu, kar povzroči zmanjšanje "temnega" toka. Amplituda toka je majhna in sorazmerna s številom absorbiranih fotonov. Včasih blisk povzroči en sam odziv, včasih dvojni odziv in včasih nobenega odziva.
Pri opičjih palicah je zmanjšanje toka kot odgovor na absorpcijo enega fotona približno 0,5 pA. To ustreza zaprtju približno 300 kanalov, tj. od 3 do 5 % vseh kanalov odprtih v temi. To dosežemo zaradi znatnega povečanja signala v presnovi kaskada cGMP. Poleg tega sta zaradi izredne stabilnosti prej omenjenih vizualnih pigmentov naključna izomerizacija in lažno zaprtje kanala zelo redka dogodka. To povzroči, da učinki posameznih svetlobnih kvantov izstopajo na ozadju zelo nizkega šuma enosmernega toka. Pokazalo se je, da električna sklopitev med fotoreceptorji zagotavlja dodaten učinek glajenja, ki zmanjša hrup v ozadju in izboljša razmerje med signalom in šumom odzivov paličice na posamezne kvante.
Literatura
1. Finn, J. T., Grunwald, M. E, in Yau, K-W. 1996. Ionski kanali, odvisni od cikličnih nukleotidov: razširjena družina z različnimi funkcijami. Annu. Rev. Physiol.58: 395-426.
2. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y, Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S. in Okada, M. 1998. Glutamatni receptorji : Delovanje možganov in prenos signalov.
VIZUALNI PIGMENTI(lat. pigmentum paint) - fotosenzitivni pigmenti fotoreceptorjev mrežnice. 3. Predmeti, ki zaznajo energijo svetlobnega impulza, opravijo zapleten fotokemični cikel. transformacije, zaradi katerih ločen vizualni receptor mrežnice, ki vsebuje 3. p (stožec ali palica), preide v vznemirjeno stanje in prenaša prejete informacije vzdolž optičnega živca do c. n. z. Kot glavni strukturni in funkcionalni del fotoreceptorske membrane vidnih celic mrežnice, 3. str. ključna vloga v mehanizmih vida (glej).
Nomenklatura in struktura vizualnih pigmentov. Vse proučevane 3. p. vretenčarjev in nevretenčarjev so kompleksi v vodi netopnega membranskega proteina opsina in z njim povezanega kromofora (retinala). Retinal ali aldehid vitamina A lahko obstaja v dveh oblikah, retinal1 in retinal2.
Po naravi kromoforja 3. je element razdeljen v dva razreda - rodopsine (glej), ki vsebujejo retinal1, in porfiropsine, ki vsebujejo retinal2. Rodopsine najdemo v mrežnici očesa vseh kopenskih in morskih živali, porfiropsine najdemo v mrežnici oči sladkovodnih živali. Nekatere ribe in dvoživke imajo 3 p., ki vsebujejo retinal hkrati! in mrežnice. Obstajajo poskusi razvrstitve 3. str na podlagi razlik v opsinih, značilnih za paličice ali stožce mrežnice. Na primer, rodopsin je kompleks retinala1 s paličastim opsinom, jodopsin je kompleks retinala1 s stožčastim opsinom, porfiropsin je retinal2 s paličastim opsinom, kompleks opsina retinal-stožček tvori cianopsin. Vendar pa je zelo težko 3. predmete razvrstiti na podlagi opsinov, saj obstaja vsaj pet različnih opsinov.
Od vseh znanih 3. predmetov so bili najbolj temeljito raziskani rodopsini, izolirani iz oči bika, žabe in lignja. Njihova prošnja. mase (mase) reda 30-40 tisoč, vsaka molekula vsebuje cca. 400 aminokislin in en kromofor. Poleg tega je oligosaharidna veriga vključena v molekulo 3. p .: 3 radikali glukozamina, 2 manoza, 1 galaktoza. Lipidi (pogl. arr. fosfolipidi) tvorijo močan kompleks z molekulo 3. str. 3. Predmeti brez lipidov ob ohranjanju svojih osnovnih spektralnih lastnosti (glej Spektralna analiza) izgubijo številne funkcionalno pomembne, na primer sposobnost obnavljanja.
Čista mrežnica ima rumena, leži maksimum njegovega absorpcijskega spektra v območju 370 nm. Opsin je brezbarven, absorpcijski maksimum je v ultravijoličnem območju (cca. 280 nm). Barva molekule rodopsina je rdečkasto rožnata, maksimalni absorpcijski spekter je cca. 500 nm. Razlog za tako močan spektralni premik med nastajanjem kompleksa (od 370 do 500 nm – tako imenovani batokromni premik) še ni dobil nedvoumne razlage.
Maksimumi absorpcijskih spektrov rodopsinov in porfiropsinov zajamejo precej široko območje vidnega spektra - od 433 do 562 nm za rodopsine in od 510 do 543 nm za porfiropsine. Če so med porfiropsine tudi 3.p stožci paglavca žabe, krapa in sladkovodne želve, torej cianopsin z maksimalnim absorpcijskim spektrom pri 620 nm, potem je to območje še širše. Razvoj mikrospektrofotometričnih metod je omogočil določitev absorpcijskih spektrov mnogih vrst posameznih fotoreceptorskih celic pri živalih in ljudeh. Glede na pridobljene podatke ima 3. stran človeške mrežnice naslednje maksimume absorpcijskega spektra: palice 498, stožci, občutljivi na modro, zeleno in rdeče - 440, 535 in 575 nm.
V nemščini se je začel študij 3. str. raziskovalec H. Muller, ki je leta 1851 opisal, kako postane rožnato-vijolična mrežnica, izvlečena iz očesa žabe, na svetlobi najprej rumenkasta, nato pa belkasta. Leta 1877 je F. Boll opisal tudi ta pojav in ugotovil, da se nekakšna snov, občutljiva na rdečo svetlobo, nahaja v vidnih celicah mrežnice in da je razbarvanje te snovi povezano z mehanizmom vida. Velike zasluge pri preučevanju 3. t. ima Kuhne (W. Kuhne, 1877), Krom je uspel identificirati 3. t. in jih podrobno preučiti. 3. p., ki ga je pridobil, je imenoval vizualno vijolična, ugotovil njegovo proteinsko naravo, raziskal nekatere njegove spektralne lastnosti in fototransformacije, odkril sposobnost 3. p., da se obnovi v temi. K študiju 3. str. biokemik in fiziolog J. Wald.
Fototransformacije vidnih pigmentov. Pod vplivom svetlobe na 3. p., se v njih zgodi fotokemični cikel. transformacije, ki temeljijo na primarni fotokemični reakciji cis-trans izomerizacije mrežnice (glej Izomerija). V tem primeru je vez med kromoforjem in proteinom motena. Zaporedje transformacij 3. p. je mogoče predstaviti na naslednji način: rodopsin (kromofor je v cis obliki) -> prelumirhodopsin -> lumirhodopsin -> metarodopsin I -> metarodopsin II -> protein opsin -> kromofor v trans obliki. Pod vplivom encima - retinol dehidrogenaze - slednji prehaja v vitamin A, ki prihaja iz zunanjih segmentov palic in stožcev v celice pigmentne plasti mrežnice. Ko je oko zatemnjeno, pride do regeneracije 3. p., za izvedbo reza je potrebna prisotnost cis-izomera vitamina A, ki služi kot začetni produkt za tvorbo kromoforja (aldehid vitamina A). potrebno. S pomanjkanjem ali odsotnostjo vitamina A v telesu je lahko tvorba rodopsina motena in se razvije kot posledica motnje vida v somraku, tako imenovani. nočna slepota (glej Hemeralopia). V procesu fototransformacije rodopsina na stopnji prehoda lumirhodopsina v metarodopsin I v receptorski celici se pojavi kot odgovor na svetlo bliskavico tako imenovanega. zgodnji (kratkolatenčni) receptorski potencial. Vendar pa ni vizualni signal, čeprav lahko služi kot eden od testov za preučevanje mehanizma transformacij 3. p v fotoreceptorski membrani. Funkcionalna vrednost je ti. poznega receptorskega potenciala je latentna doba to-rogo (5-10 msec) sorazmerna s časom nastanka metarodopsina II. Predpostavlja se, da reakcija prehoda metarodopsina I v metarodopsin II zagotavlja pojav vizualnega signala.
3. Ker se predmeti na svetlobi nenehno obarvajo, morajo obstajati mehanizmi za njihovo stalno obnavljanje. Nekatere med njimi so izjemno hitre (fotoregeneracija), druge precej hitre (biokemične, regeneracijska, temna), tretje počasne (sinteza 3. p. Med stalnim obnavljanjem fotoreceptorske membrane v vidni celici). Fotoregeneracija ima fiziolno vrednost pri nevretenčarjih (na primer pri glavonožci- lignji, hobotnice). V mehanizmu biokem. regeneracija 3. str pri vretenčarjih očitno igra pomembno vlogo encim izomeraza (glej), ki zagotavlja izomerizacijo trans-retinala (ali trans-vitamina A) ponovno v cis izomerno obliko. Vendar dokončnih dokazov o obstoju takega encima ni. Podobna reakcija tvorbe molekule 3. p v prisotnosti 11-cis-izomera retinala in opsina v sistemu poteka enostavno, brez porabe energije. Najdena je sposobnost beljenega rodopsina za reakcijo fosforilacije (glej); domneva se, da je ta reakcija ena od povezav v mehanizmu svetlobne prilagoditve vidne celice.
Bibliografija: Ackerman Yu. Biofizika, trans. iz angleščine, M., 1964; Willie K. in Det e V. Biologija, prev. iz angleščine, M., 1974, bibliografija; Konev S. V. in Volotovski I. D. Uvod v molekularno fotobiologijo, str. 61, Minsk, 1971; Ostrovsky M. A. in Fedorovich I. B. Fotoinducirane spremembe v fotoreceptorski membrani, v knjigi: Struktura in funkcije biol, membran, ed. A. S. Troshina in drugi, str. 224, M., 1975, bibliogr.; Fiziologija senzoričnih sistemov, ed. G. V. Gershuni, 1. del, str. 88, L., 1971; Biokemija in fiziologija vidnih pigmentov, ed. od H. Langer, B. a. o., 1973; Priročnik senzorične fiziologije, ed. od H. A. R. Jung a. o., v. 7, pt 1-2, B., 1972.
M. A. Ostrovskega.