Vizualni pigmenti. Rudimentarna zona epidermisa kože sastoji se od slojeva. Dnevni i noćni vid
Vidni pigmenti koncentrirani su u membranama vanjskih segmenata. Svaki štapić sadrži oko 10 8 molekula pigmenta. Organizirani su u nekoliko stotina diskretnih diskova (oko 750 u majmunskom štapu) koji nisu povezani s vanjskom membranom. U čunjićima se pigment nalazi u posebnim pigmentnim naborima, koji su nastavak vanjske stanične membrane fotoreceptora. Molekule pigmenta čine oko 80% svih proteina diska. Vidni pigmenti toliko su gusto zbijeni u membranama vanjskog segmenta da razmak između dviju molekula vidnog pigmenta u štapiću ne prelazi 10 nm. Ovo gusto pakiranje povećava vjerojatnost da će foton svjetlosti koji prolazi kroz sloj fotoreceptorskih stanica biti uhvaćen. Postavlja se sljedeće pitanje: kako nastaju signali kada svjetlo apsorbiraju vizualni pigmenti?
Apsorpcija svjetlosti vizualnim pigmentimaPsihofiziološkim, biokemijskim i molekularnim tehnikama proučavani su događaji koji se zbivaju kada svjetlo apsorbira pigment štapića, rodopsin. Molekula vidnog pigmenta sastoji se od dvije komponente: proteina zvanog opsin i kromofora, 11-cis-vitamin A aldehida zvanog retinal (Slika 1). Treba pojasniti da kromofor sadrži kemijsku skupinu koja daje boju spoju. Kvantitativna svojstva apsorpcijske sposobnosti pigmenata proučavana su spektrofotometrijski. Kad se rodopsin, vidni pigment štapića, osvijetli svjetlom različitih valnih duljina, najbolje se apsorbira plavozeleno svjetlo valne duljine od oko 500 nm. Sličan rezultat dobiven je kada je zasebna šipka osvijetljena pod mikroskopom snopovima svjetlosti različitih valnih duljina. Bio identificiran zanimljiva ovisnost između apsorpcijskog spektra rodopsina i naše percepcije svjetla u sumrak. Kvantitativne psihofizičke studije provedene na ljudima pokazale su da je plavkasto-zelena dnevna svjetlost valne duljine od oko 500 nm optimalna za percepciju sumračne svjetlosti u mraku. Tijekom dana, kada su štapići neaktivni i koriste se samo čunjići, najosjetljiviji smo na crvenu boju koja odgovara apsorpcijskom spektru čunjića (o tome ćemo kasnije).
Kada rodopsin apsorbira jedan foton, retinal prolazi kroz fotoizomerizaciju i prelazi iz 11-cis u trans konfiguraciju. Ovaj prijelaz se događa vrlo brzo: za oko 10-12 sekundi. Nakon toga, proteinski dio pigmenta također prolazi kroz niz transformacijskih promjena, uz stvaranje niza međuproizvoda. Jedna konformacija proteinskog dijela, metarodopsin II, najvažniji je za prijenos signala (o tome ćemo raspravljati kasnije u ovom poglavlju). Slika 2 prikazuje slijed događaja tijekom izbjeljivanja i regeneracije aktivnog rodopsina. Metarhodopsin II nastaje unutar 1 ms. Regeneracija pigmenta nakon njegovog raspadanja odvija se polako, unutar nekoliko minuta; To zahtijeva transport retine od fotoreceptora do pigmentnog epitela.
Struktura rodopsinaNa molekularnoj razini, protein opsin sastoji se od 348 aminokiselinskih ostataka, koji tvore 7 hidrofobnih zona, od kojih se svaka sastoji od 20-25 aminokiselina, čineći 7 transmembranskih spirala. N-kraj molekule nalazi se u izvanstaničnom prostoru (tj. unutar diska štapića), a C-kraj se nalazi u citoplazmi.
U mraku je 11-cis-retinal čvrsto vezan za protein opsin. Hvatanje fotona rezultira izomerizacijom cijelog cis retinala u tron retinala. U tom slučaju kompleks opsin all-trons-retinal brzo prelazi u metarodo psin II, koji se disocira na opsin i all-trons retinal. Regeneracija rodopsina ovisi o interakciji fotoreceptora i pigmentnih epitelnih stanica. Metarhodopsin II uključuje i održava sekundarni glasnički sustav u aktivnom stanju.
Retinal je povezan s opsinom preko lizinskog ostatka koji se nalazi u sedmom transmembranskom segmentu. Opsin pripada obitelji proteina sa 7 transmembranskih domena, koja također uključuje metabotropne neurotransmiterske receptore, kao što su adrenergički i muskarinski receptori. Poput rodopsina, ovi receptori prenose signale sekundarnim glasnicima putem aktivacije G proteina. Rhodopsin je iznenađujuće stabilan u mraku. Bayor je izračunao da je za spontanu toplinsku izomerizaciju molekule rodopsina potrebno oko 3000 godina, ili 10 23 više od fotoizomerizacije.
Vizualni pigmenti
Vizualni pigmenti
Struktura rodopsina
Čunjići i vid u boji
Daltonizam
Književnost
Vizualni pigmenti
Vidni pigmenti koncentrirani su u membranama vanjskih segmenata. Svaki štapić sadrži oko 10 8 molekula pigmenta. Organizirani su u nekoliko stotina diskretnih diskova (oko 750 u majmunskom štapu) koji nisu povezani s vanjskom membranom. U čunjićima se pigment nalazi u posebnim pigmentnim naborima, koji su nastavak vanjske stanične membrane fotoreceptora. Molekule pigmenta čine oko 80% svih proteina diska. Vidni pigmenti toliko su gusto zbijeni u membranama vanjskog segmenta da razmak između dviju molekula vidnog pigmenta u štapiću ne prelazi 10 nm. Ovo gusto pakiranje povećava vjerojatnost da će foton svjetlosti koji prolazi kroz sloj fotoreceptorskih stanica biti uhvaćen. Postavlja se sljedeće pitanje: kako nastaju signali kada svjetlo apsorbiraju vizualni pigmenti?
Apsorpcija svjetlosti vizualnim pigmentima
Psihofiziološkim, biokemijskim i molekularnim tehnikama proučavani su događaji koji se zbivaju kada svjetlo apsorbira pigment štapića, rodopsin. Molekula vidnog pigmenta sastoji se od dvije komponente: proteina zvanog opsin i kromofora, 11-cis-vitamin A aldehida zvanog retinal (Slika 1). Treba pojasniti da kromofor sadrži kemijsku skupinu koja daje boju spoju. Kvantitativna svojstva apsorpcijske sposobnosti pigmenata proučavana su spektrofotometrijski. Kad se rodopsin, vidni pigment štapića, osvijetli svjetlom različitih valnih duljina, najbolje se apsorbira plavozeleno svjetlo valne duljine od oko 500 nm. Sličan rezultat dobiven je kada je zasebna šipka osvijetljena pod mikroskopom snopovima svjetlosti različitih valnih duljina. Identificiran je zanimljiv odnos između apsorpcijskog spektra rodopsina i naše percepcije svjetla u sumrak. Kvantitativne psihofizičke studije provedene na ljudima pokazale su da je plavkasto-zelena dnevna svjetlost valne duljine od oko 500 nm optimalna za percepciju sumračne svjetlosti u mraku. Tijekom dana, kada su štapići neaktivni i koriste se samo čunjići, najosjetljiviji smo na crvenu boju koja odgovara apsorpcijskom spektru čunjića (o tome ćemo kasnije).
Kada rodopsin apsorbira jedan foton, retinal prolazi kroz fotoizomerizaciju i prelazi iz 11-cis u trans konfiguraciju. Ovaj prijelaz se događa vrlo brzo: za oko 10-12 sekundi. Nakon toga, proteinski dio pigmenta također prolazi kroz niz transformacijskih promjena, uz stvaranje niza međuproizvoda. Jedna konformacija proteinskog dijela, metarodopsin II, najvažniji je za prijenos signala (o tome ćemo raspravljati kasnije u ovom poglavlju). Slika 2 prikazuje slijed događaja tijekom izbjeljivanja i regeneracije aktivnog rodopsina. Metarhodopsin II nastaje unutar 1 ms. Regeneracija pigmenta nakon njegovog raspadanja odvija se polako, unutar nekoliko minuta; To zahtijeva transport retine od fotoreceptora do pigmentnog epitela.
Struktura rodopsina
Na molekularnoj razini, protein opsin sastoji se od 348 aminokiselinskih ostataka, koji tvore 7 hidrofobnih zona, od kojih se svaka sastoji od 20-25 aminokiselina, čineći 7 transmembranskih spirala. N-kraj molekule nalazi se u izvanstaničnom prostoru (tj. unutar diska štapića), a C-kraj se nalazi u citoplazmi.
Sl. 1. Struktura rodopsina kralježnjaka ugrađenog u fotoreceptorsku membranu. Zavojnica se lagano zakreće kako bi se pokazalo mjesto retine (označeno crnom bojom). C - C-kraj, N - N-kraj.
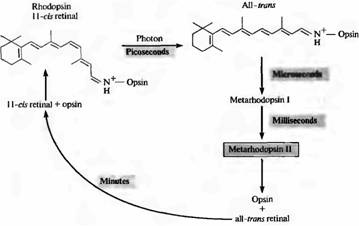
sl.2. Blijeđenje rodopsina na svjetlu. U mraku je 11-cis-retinal čvrsto vezan za protein opsin. Hvatanje fotona rezultira izomerizacijom cijelog cis retinala u tron retinala. U tom slučaju kompleks opsin all-trons-retinal brzo prelazi u metarodo psin II, koji se disocira na opsin i all-trons retinal. Regeneracija rodopsina ovisi o interakciji fotoreceptora i pigmentnih epitelnih stanica. Metarhodopsin II uključuje i održava sekundarni glasnički sustav u aktivnom stanju.
Retinal je povezan s opsinom preko lizinskog ostatka koji se nalazi u sedmom transmembranskom segmentu. Opsin pripada obitelji proteina sa 7 transmembranskih domena, koja također uključuje metabotropne neurotransmiterske receptore, kao što su adrenergički i muskarinski receptori. Poput rodopsina, ovi receptori prenose signale sekundarnim glasnicima putem aktivacije G proteina. Rhodopsin je iznenađujuće stabilan u mraku. Bayor je izračunao da je za spontanu toplinsku izomerizaciju molekule rodopsina potrebno oko 3000 godina, ili 10 23 više od fotoizomerizacije.
Čunjići i vid u boji
Nevjerojatna istraživanja i eksperimenti koje su izveli Young i Helmholtz u 19. stoljeću skrenuli su pozornost na vrlo važno pitanje o kolornom vidu, a sami znanstvenici dali su jasno i precizno objašnjenje ovog fenomena. Njihov zaključak o postojanju triju različite vrste fotoreceptora u boji izdržao je test vremena i naknadno je potvrđen na molekularnoj razini. Opet možemo citirati Helmholtza koji je usporedio percepciju svjetla i zvuka, boje i zvučnog tona. Može se pozavidjeti na jasnoći, snazi i ljepoti njegove misli, posebno u usporedbi sa zbunjujućim vitalističkim konceptima koji su bili rašireni u 19. stoljeću:
Sve razlike u tonovima boja ovise o kombinaciji u različitim omjerima triju primarnih boja... crvene, zelene i ljubičaste... Baš kao što percepcija sunčeve svjetlosti i njezine topline ovisi... o tome padaju li sunčeve zrake na živaca, koji dolaze od receptora vida ili od receptora toplinske osjetljivosti. Kao što je Young sugerirao u svojoj hipotezi, razlika u percepciji različitih boja jednostavno ovisi o tome koji od 3 tipa fotoreceptora je više aktiviran tom svjetlošću. Kad su sve tri vrste in jednako uzbuđeno, ispada bijelo...
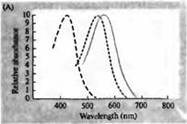
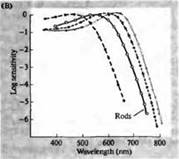
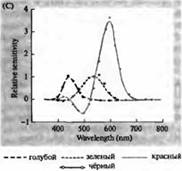
Riža. 3. Spektri osjetljivosti ljudskih fotoreceptora i razni vizualni pigmenti. (A) Krivulje spektra osjetljivosti triju vizualnih pigmenata u boji, koje pokazuju vrhunce apsorpcije na valnim duljinama koje odgovaraju plavoj, zelenoj i crvenoj boji. (B) Spektri osjetljivosti čunjića na plavu, zelenu i crvenu boju i štapića (prikazano crno) kod makakija. Odgovori su zabilježeni korištenjem usisnih elektroda, usrednjeni i normalizirani. Krivulje štapićastog spektra dobivene su proučavanjem vizualnih pigmenata kod ljudi. (C) Usporedba spektra čunjića majmuna i čovjeka pomoću testa osjetljivosti na boju. Kontinuirana krivulja prikazuje eksperiment za određivanje osjetljivosti na boje kod ljudi prikazivanjem svjetla različitih valnih duljina. Isprekidana linija prikazuje rezultate predviđene iz snimanja struja u pojedinačnim čunjevima, nakon korekcije za apsorpciju svjetla u leći i pigmentima na putu do vanjskog segmenta. Slaganje između rezultata oba eksperimenta je iznenađujuće visoko.
Projiciramo li dvije zrake svjetlosti različitih boja istovremeno na bijeli ekran... vidimo samo jednu boju, više-manje različitu od obje ove boje. Nevjerojatnu činjenicu da smo sposobni zamijetiti sve nijanse u sastavu vanjske svjetlosti pomoću mješavine tri osnovne boje možemo bolje razumjeti ako usporedimo suho oko... U slučaju zvuka... čujemo duži valovi kao niski tonovi, a kraći valovi - kao visoki i prodorni, osim toga, uho je sposobno uhvatiti više zvučnih valova u isto vrijeme, tj. puno bilješki. Međutim, u ovom slučaju one se ne stapaju u jedan složeni akord, kao što se različite boje... stapaju u jednu složenu boju. Oko ne može uočiti razliku ako zamijenimo narančasta boja do crvene ili žute; ali ako čujemo da note C i E zvuče istovremeno, takav zvuk nam se ne čini kao D nota. Kad bi uho percipiralo glazbene tonove na isti način na koji oko percipira boje, svaki bi akord mogao biti predstavljen kombinacijom tri stalne note, jedne vrlo niske, jedne vrlo visoke i jedne između, uzrokujući sve moguće glazbene efekte samo promjenama u relativna glasnoća ove tri note... Međutim, mi smo u stanju vidjeti gladak prijelaz boja iz jedne u drugu kroz beskonačan broj nijansi i gradacija... Način na koji percipiramo svaku od boja... ovisi uglavnom na strukturu našeg živčani sustav. Mora se priznati da trenutno nije opisana anatomska osnova za potvrdu teorije percepcije boja ni kod ljudi ni kod četveronožaca.
Ova točna i dalekovidna predviđanja potvrđena su nizom različitih opažanja. Koristeći spektrofotometriju, Wald, Brauk, McNicol i Dartnall i njihovi kolege pokazali su prisutnost tri vrste čunjića s različitim pigmentima u ljudskoj mrežnici. Baylor i njegovi kolege također su uspjeli skrenuti struju iz čunjeva majmuna i ljudi. Utvrđeno je da tri populacije čunjića imaju različite, ali preklapajuće raspone osjetljivosti na plavi, zeleni i crveni dio spektra. Optimalne valne duljine za uzbudljive električne signale točno su se poklapale s vrhovima apsorpcije svjetlosti vizualnih pigmenata, utvrđenih spektrofotometrijom i psihofizičkim eksperimentima mjerenjem osjetljivosti oka na spektar boja. U konačnici, Natais je klonirao i sekvencirao gene koji kodiraju pigment opsin u tri vrste čunjića koji su osjetljivi na crveni, zeleni i plavi spektar.
Kako to da molekule različitih vizualnih pigmenata mogu preferencijalno uhvatiti svjetlost određene valne duljine? Ispostavilo se da je rodopsin vidni pigment štapića i da sva tri vidna pigmenta čunjića sadrže isti kromofor, 11-cis-retinal. Međutim, aminokiselinske sekvence proteinskog dijela pigmenta međusobno se razlikuju. Razlike u samo nekoliko aminokiselina objašnjavaju njihovu različitu osjetljivost na spektar.
Daltonizam
Iako jedna vrsta fotoreceptora nije sposobna sama percipirati boju, tri vrste čunjića, kao što je prikazano na sl. 4 su već sposobni.
U principu, dvije vrste čunjića s različitim pigmentima bile bi dovoljne za raspoznavanje boja, ali u ovom slučaju bi se više kombinacija valnih duljina percipiralo jednako. Slična situacija nastaje kada osoba pati od sljepoće za boje. Takvi ljudi, kako je pokazao Nathans, imaju genetske defekte koji dovode do odsutnosti jednog od pigmenata. S visine trenutnog stanja znanosti, ne možemo a da se ne začudimo kako lijepo molekularni mehanizmi potvrđuju briljantno i iznenađujuće točno razmišljanje Younga i Helmholtza.
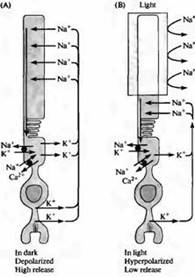
Riža. 4. "Tamna" struja u šipki. (A) U mraku, natrijevi ioni prolaze kroz kationske kanale u vanjskom segmentu štapića, uzrokujući depolarizaciju; Ioni kalcija također mogu proći kroz te kationske kanale. Strujna petlja prolazi kroz prevlaku štapića zbog činjenice da struja kalija teče u smjeru prema van u unutarnjem segmentu membrane. (B) Kada dođe do osvjetljenja vanjskog segmenta, kanali se zatvaraju zbog smanjenja intracelularne koncentracije cGMP, a štapić se hiperpolarizira. Hiperpolarizacija dovodi do smanjenja otpuštanja transmitera. Koncentracije natrija, kalija i kalcija održavaju se unutar štapića pomoću posebnih pumpi i ionskih izmjenjivača koji se nalaze u području unutarnjeg segmenta (crni krugovi). U vanjskom segmentu nalaze se i transporteri kalcija.
Njihova ideja da se osnovni atributi vida boja i sljepoće za boje nalaze u samim fotoreceptorima potvrđena je izravnim fiziološkim mjerenjima, kao i proučavanjem razlika u strukturi pigmenata na genetskoj i proteinskoj razini, potencijal je na redu od -40 mV, što je daleko od ravnotežnog kalijevog potencijala E K, koji iznosi 80 mV.

Riža. 5. Uloga cGMP-a u otvaranju natrijevih kanala u vanjskom segmentu štapića. Aktivnost pojedinačnih kanala zabilježena je korištenjem unutarnje vanjske stezaljke smještene u otopini s različitim koncentracijama cGMP-a. Otvaranje kanala dovodi do skretanja struje prema gore. Učestalost otvaranja kanala bila je vrlo niska u kontroli, a dodavanje cGMP-a dovelo je do povećanja učestalosti otvaranja pojedinačnih kanala, što je izravno ovisilo o koncentraciji cGMP-a.
Dolaznu struju u mraku nose primarno natrijevi ioni slijedeći elektrokemijski gradijent kroz kationske kanale vanjskog segmenta. Hiperpolarizacija fotoreceptora pod utjecajem svjetlosti uzrokovana je zatvaranjem ovih kanala, što dovodi do pomaka membranskog potencijala prema E K.
Svojstva fotoreceptorskih kanala
Kationski kanali vanjskog segmenta pod fiziološkim uvjetima imaju omjer vodljivosti kalcij/natrij/kalij iona od 12,5:1,0:0,7 i vodljivost pojedinačnog kanala od oko 0,1 pS 21). Budući da je koncentracija natrija mnogo viša od koncentracije kalcija, oko 85% ulazne struje dolazi od natrijevih iona. Pogonska sila iona kalija usmjerena je prema van stanice. Kada ioni kalcija prolaze kroz kanal, ti se ioni čvrsto vežu za stijenku pore i tako ometaju prolaz drugih iona. Zbog toga uklanjanje kalcija iz izvanstaničnog okoliša dovodi do lakšeg prolaska kalijevih i natrijevih iona kroz kanale, čija se vodljivost povećava na 25 pS.
Fesenko, Yau, Baylor, Strayer i suradnici pokazali su da ciklički GMP igra ulogu unutarnjeg prijenosnika signala od diska do površine membrane. Kao što je prikazano na slici 4, visoka koncentracija cGMP u citoplazmi održava kationske kanale otvorenima. Kako koncentracija cGMP-a opada s unutarnje površine membrane, otvaranje kationskih kanala postaje rijedak događaj. Dakle, membranski potencijal fotoreceptora odraz je koncentracije cGMP-a u citoplazmi: što je veća koncentracija cGMP-a, veća je depolarizacija stanice. Koncentracija cGMP-a pak ovisi o intenzitetu upadne svjetlosti. Povećanje intenziteta svjetla dovodi do smanjenja koncentracije cGMP i smanjuje udio otvorenih kanala. U nedostatku cGMP-a, gotovo svi kanali su zatvoreni, a otpor membrane vanjskog segmenta približava se otporu lipidnog dvosloja.
Molekularna struktura cGMP-uključenih kanala
Izolirana je cDNA za ionske kanale vanjskog segmenta štapića, a aminokiselinske sekvence podjedinica tih kanala određene su u retini ljudi, goveda, miševa i kokoši. Pronađena je značajna sličnost DNK za ove i druge cGMP-om upravljane ionske kanale - poput onih koji se nalaze u olfaktornom sustavu
Prijenos signala u fotoreceptorima
Kako fotoizomerizacija rodopsina dovodi do promjena u membranskom potencijalu? Dugi niz godina smatralo se da je neka vrsta unutarnjeg glasnika potrebna za pobuđivanje električnih signala u štapićima i čunjićima. Jedan od razloga za sumnju da se informacija o apsorpciji fotona u području vanjskog segmenta štapića prenosi pomoću nosača bila je činjenica da se sam pigment rodopsin nalazi unutar diska, a signal se širi kroz citoplazmu do vanjsku membranu. Drugi razlog bio je značajan porast odaziva. Baylor i kolege, proučavajući fotoreceptore kornjača, pokazali su da se smanjenje vodljivosti membrane i zabilježenih električnih signala događa čak i nakon apsorpcije samo jednog fotona i aktivacije jedne od 10 8 molekula pigmenta.
Redoslijed događaja u kojima aktivirana molekula fotopigmenta mijenja potencijal membrane razjašnjen je proučavanjem vanjskih segmenata štapića i čunjića pomoću stezanja potencijala s dvije elektrode i tehnikama molekularne biologije. Dijagram prijenosa signala od apsorpcije fotona svjetlosti do električnog signala prikazan je na sl. 3.
U mraku, stalna ulazna "tamna" struja teče u vanjskim segmentima štapića i čunjića. Kao rezultat njihovog membranskog sustava. Membranska područja ovih kanala pokazuju strukturne sličnosti s drugim kation-selektivnim kanalima, posebno u S4 regiji i u regiji koja tvori ionsku poru. Fotoreceptorski ionski kanali su tetrameri sastavljeni od najmanje 2 različite proteinske podjedinice s molekulskom masom od 63 odnosno 240 kDa.
Intracelularna mjesta vezanja nukleotida nalaze se blizu karboksilnog kraja podjedinica. Ekspresija ovih podjedinica u oocitima rezultira stvaranjem kationskih kanala koji imaju svojstva slična onima koja se nalaze u vanjskim segmentima štapića: aktivira ih cGMP i imaju očekivane omjere vodljivosti i propusnosti.

Riža. 6. Mehanizam aktivacije G-proteina tijekom aktivacije molekule fotopigmenta. G-protein transducin veže GTP u prisutnosti metarodopsina II, što dovodi do aktivacije fosfodiesteraze, koja pak hidrolizira cGMP. Kako koncentracija cGMP pada, natrijevi kanali se zatvaraju.
Ciklička GMP metabolička kaskada
Lanac događaja koji dovode do smanjenja koncentracije cGMP-a i kasnijeg zatvaranja ionskih kanala prikazan je na slici 5. Smanjenje intracelularne koncentracije cGMP-a uzrokovano je svjetlom, što dovodi do stvaranja metarodopsina II, intermedijera u razgradnji fotopigmenta. Metharhodopsin II djeluje, pak, na G-protein transducin, koji se sastoji od 3 polipeptidna lanca
Interakcija metarodopsina II i transducina dovodi do zamjene molekule GDP vezane na G-protein GTP-om. Ovo aktivira podjedinicu G proteina, koja se odvaja od podjedinica i zauzvrat aktivira fosfodiesterazu smještenu u području membrane: enzim koji hidrolizira cGMP. Koncentracija cGMP-a opada, ima manje otvorenih ionskih kanala, a štapić se hiperpolarizira. Kaskada je prekinuta fosforilacijom C-kraja aktivnog metarodopsina II. Ključna uloga cGMP-a u regulaciji stanja kationskih kanala potvrđena je biokemijskim pokusima. Osvjetljenje fotoreceptora može uzrokovati pad razine cGMP-a u stanici za 20%.
Receptori kralježnjaka koji se depolariziraju kada su izloženi svjetlu
Zanimljiva iznimka gore navedenom mehanizmu fotorecepcije su neki receptori kralježnjaka. Gušteri imaju treće oko smješteno na vrhu glave. Sadrži male "čunjeve" koji su sposobni percipirati sliku sličnu onoj koju percipiraju glavne (bočne) oči. Ti su fotoreceptori, međutim, poznati po tome što se depolariziraju kad su osvijetljeni. Kanali kojima upravljaju nukleotidi ovdje imaju sličnu strukturu i funkciju fotoreceptorima drugih kralježnjaka, uz jednu iznimku: aktivacija fotoreceptora i G proteina dovodi do povećanja koncentracije cGMP. Kao rezultat toga, kanali vanjskog segmenta se otvaraju i kationi se usmjeravaju u stanicu, tvoreći "svjetlosnu" struju. To se događa zbog inhibicije aktivnosti fosfodiesteraze u mraku. Zbog toga se u oku guštera događa sljedeći slijed događaja: svjetlost - [cGMP] - otvaranje kationskih kanala vanjskog segmenta - depolarizacija.
Pojačanje signala u cGMP kaskadi
Dvostupanjska cGMP kaskada osigurava značajno povećanje početnog signala, što objašnjava izuzetnu osjetljivost štapića na svjetlost. Prvo, jedna molekula aktivnog metarodopsina II katalizira dodavanje mnogih GTP molekula umjesto GDP-a i tako oslobađa stotine podjedinica G proteina. Drugo, svaka podjedinica aktivira jednu molekulu fosfodiesteraze na disku, koja je sposobna razgraditi ogroman broj cGMP molekula koje se nalaze u citoplazmi i tako dovesti do zatvaranja velikog broja kanala.
Signali kao odgovor na pojedinačne kvante svjetlosti
Potaknuli su dokazi da pojedinačni kvanti svjetlosti mogu proizvesti percipirani osjet svjetlosti veliki broj pitanja. Koliki je ovaj pojedinačni odgovor? Kako se taj signal odvaja od razine šuma? I kako se takve informacije pouzdano prenose od mrežnice do viših vidnih centara? Kako bi izmjerili signale kao odgovor na pojedinačne kvante svjetlosti, Baylor i njegovi kolege zabilježili su struje iz pojedinačnih štapića u mrežnici žaba krastača, majmuna i ljudi. Ovi pokusi predstavljaju jedinstven primjer pokusa koji pokazuje kako takav težak proces, kao i percepcija slabih bljeskova svjetlosti, može biti u korelaciji s promjenama koje se događaju na razini pojedinačnih molekula.
Postupak izolacije retinalnog fragmenta iz životinje ili kadaverskog materijala mora se provesti u mraku za ove pokuse. Za mjerenje struje, vanjski segment šipke se usisava u tanku pipetu. Kao što se i očekivalo, ovi eksperimenti pokazuju da u mraku struja stalno teče u vanjski segment. Bljeskovi svjetla dovode do zatvaranja kanala u vanjskom segmentu, uzrokujući smanjenje "tamne" struje. Amplituda struje je mala i proporcionalna broju apsorbiranih kvanta. Ponekad bljeskalica proizvede jedan odgovor, ponekad dvostruki odgovor, a ponekad uopće ne odgovori.
Kod majmunskih štapića, smanjenje struje kao odgovor na apsorpciju jednog fotona je oko 0,5 pA. To odgovara zatvaranju oko 300 kanala, tj. od 3 do 5% svih kanala otvoreno je u mraku. To se postiže zahvaljujući značajnom povećanju signala u metabolizmu cGMP kaskada. Štoviše, zbog ekstremne stabilnosti vizualnih pigmenata spomenutih ranije, nasumična izomerizacija i lažno zatvaranje kanala vrlo su rijetki događaji. To uzrokuje da se učinci pojedinačnih svjetlosnih kvanta ističu na pozadini vrlo niske konstantne buke. Pokazalo se da električno spajanje kroz praznine između fotoreceptora daje dodatni učinak izglađivanja koji smanjuje pozadinsku buku i poboljšava omjer signala i buke odgovora štapića na pojedinačne kvante.
Književnost
1. Finn, J. T., Grunwald, M. E, i Yau, K-W. 1996. Ionski kanali upravljani cikličkim nukleotidima: Proširena obitelj s različitim funkcijama. Annu. vlč. Physiol.58: 395-426.
2. Nakanishi, S., Nakajima, Y., Masu, M., Ueda, Y., Nakahara, K., Watanabe, D., Yamaguchi, S., Kawabata, S., i Okada, M. 1998. Glutamat receptori : Funkcija mozga i prijenos signala.
VIZUALNI PIGMENTI(lat. pigmentum boja) - fotoosjetljivi pigmenti fotoreceptora mrežnice. Opažajući energiju svjetlosnog pulsa, 3. predmeti prolaze složeni fotokemijski ciklus. transformacije, zbog čega zasebni vizualni receptor mrežnice, koji sadrži 3. p. (konus ili šipka), prelazi u uzbuđeno stanje i prenosi primljene informacije u središnji živčani sustav duž optičkog živca. n. S. Budući da je glavni strukturni i funkcionalni dio fotoreceptorske membrane vidnih stanica mrežnice, 3. str. glavna uloga u mehanizmima vida (vidi).
Nomenklatura i struktura vizualnih pigmenata. Svi proučavani 3.p. kralješnjaka i beskralješnjaka su kompleksi membranskog proteina opsina netopivog u vodi i pripadajućeg kromofora (retinala). Retinal ili aldehid vitamina A može postojati u dva oblika - retinal1 i retinal2.
Prema prirodi kromofora, 3. p. podijeljeni su u dvije klase - rodopsine (vidi), koji sadrže retinal1, i porfiropsine, koji sadrže retinal2. Rodopsini se nalaze u mrežnici očiju svih kopnenih i morskih životinja, porfiropsini se nalaze u mrežnici očiju slatkovodnih životinja. Kod nekih riba i vodozemaca nađeni su 3. predmeti koji istovremeno sadrže retinal! i mrežnice. Postoje pokušaji klasificiranja 3. p. na temelju razlika u opsinima specifičnim za štapiće ili čunjiće retine. Na primjer, rodopsin je kompleks retinal1 sa štapićastim opsinom, jodopsin je kompleks retinal1 sa konusnim opsinom, porfiropsin je retinal2 kompleks sa štapićastim opsinom, retinal-konusni opsin kompleks tvori cijanopsin. Međutim, vrlo je teško klasificirati 3. p. na temelju opsina, budući da postoji najmanje pet različitih opsina.
Od svih poznatih 3. p. najpotpunije su proučeni rodopsini izolirani iz očiju bikova, žaba i lignji. Oni kažu težina (masa) je oko 30-40 tisuća, svaka molekula sadrži cca. 400 aminokiselina i jedan kromofor. Osim toga, molekula 3. p. uključuje oligosaharidni lanac: 3 glukozaminska radikala, 2 manoza, 1 galaktoza. Lipidi (uglavnom fosfolipidi) tvore jak kompleks s molekulom 3.p. Zadržavajući svoja osnovna spektralna svojstva (vidi Spektralna analiza), 3.p bez lipida gube niz funkcionalno važnih, na primjer, sposobnost oporavka.
Čista mrežnica ima žuta boja, maksimum njegovog apsorpcijskog spektra leži u području od 370 nm. Opsin je bezbojan, maksimum apsorpcije je u ultraljubičastom području (cca. 280 nm). Boja molekule rodopsina je crvenkasto-ružičasta, maksimalni apsorpcijski spektar je cca. 500 nm. Razlog tako jakog spektralnog pomaka tijekom formiranja kompleksa (od 370 do 500 nm – tzv. batokromski pomak) još nije dobio jednoznačno objašnjenje.
Maksimalni apsorpcijski spektri rodopsina i porfiropsina pokrivaju prilično široko područje vidljivog spektra - od 433 do 562 nm za rodopsine i od 510 do 543 nm za porfiropsine. Ako se 3. čunjevi punoglavca žabe, šarana i slatkovodne kornjače također smatraju porfiropsinima, tj. cijanopsinima s maksimalnim apsorpcijskim spektrom na 620 nm, onda se ovo područje pokazuje još širim. Razvoj mikrospektrofotometrijskih metoda omogućio je određivanje apsorpcijskih spektara mnogih vrsta pojedinačnih fotoreceptorskih stanica kod životinja i ljudi. Prema dobivenim podacima, 3. p. ljudske mrežnice imaju sljedeće maksimume apsorpcijskog spektra: štapići 498, čunjići osjetljivi na plavo, zeleno i crveno - 440, 535 i 575 nm, redom.
Na njemačkom jeziku započeo je studij 3. str. istraživač H. Muller, koji je 1851. opisao kako ružičasto-ljubičasta mrežnica, izvađena iz oka žabe, postaje žućkasta, a zatim bjelkasta na svjetlu. Godine 1877. F. Boll je također opisao ovaj fenomen, zaključivši da u vidnim stanicama mrežnice postoji neka vrsta tvari osjetljive na crvenu svjetlost i da je promjena boje te tvari povezana s mehanizmom vida. Velike zasluge za proučavanje 3. točke pripadaju Kuhneu (W. Kuhne, 1877), koji je uspio izdvojiti 3. točke i detaljno ih proučiti. Nazvao je 3.p. kojeg je ekstrahirao vizualno ljubičastom, utvrdio njegovu proteinsku prirodu, istražio neka od njegovih spektralnih svojstava i fototransformacija i otkrio sposobnost 3.p.-a da se obnavlja u mraku. Veliki doprinos proučavanju 3. str. biokemičar i fiziolog J. Wald.
Fototransformacije vidnih pigmenata. Kada se 3.p. izloži svjetlu, u njima se događa fotokemijski ciklus. transformacije, koje se temelje na primarnoj fotokemijskoj reakciji cis-trans izomerizacije retinala (vidi Izomerija). U tom slučaju dolazi do prekida veze između kromofora i proteina. Redoslijed transformacija 3. p. može se prikazati na sljedeći način: rodopsin (kromofor je u cis obliku) -> prelumirodopsin -> lumirhodopsin -> metarhodopsin I -> metarhodopsin II -> protein opsin -> kromofor u trans obliku. Pod utjecajem enzima - retinol dehidrogenaze - potonji se pretvara u vitamin A, koji dolazi iz vanjskih segmenata štapića i čunjića u stanice pigmentnog sloja mrežnice. Kada oko potamni, dolazi do regeneracije 3. p., a za provedbu reza potrebna je prisutnost cis-izomera vitamina A, koji služi kao polazni produkt za stvaranje kromofora (vitamin A aldehid). U slučaju manjka ili odsustva vitamina A u organizmu, može doći do poremećaja stvaranja rodopsina i kao posljedica toga može se razviti poremećaj vida u sumrak, tzv. noćno sljepilo (vidi Hemeralopia). U procesu fototransformacija rodopsina u fazi prijelaza lumirhodopsina u metarodopsin I u receptorskoj stanici javlja se kao odgovor na svijetli bljesak tzv. rani (kratkolatentni) receptorski potencijal. Međutim, on nije vizualni signal, iako može poslužiti kao jedan od testova za proučavanje mehanizma transformacija 3. p. u fotoreceptorskoj membrani. Funkcionalni značaj ima tzv. kasni receptorski potencijal, čije je latentno razdoblje (5-10 ms) usporedivo s vremenom nastanka metarodopsina II. Pretpostavlja se da reakcija prijelaza metarodopsina I u metarodopsin II osigurava pojavu vizualnog signala.
Budući da 3. p. stalno mijenjaju boju na svjetlu, moraju postojati mehanizmi za njihovu stalnu obnovu. Neki od njih su izuzetno brzi (fotoregeneracija), drugi su prilično brzi (biokemijski, regeneracija, Dark), treći su spori (sinteza 3. p. tijekom stalnog obnavljanja fotoreceptorske membrane u vidnoj stanici). Fotoregeneracija je fiziol, važan kod beskralježnjaka (npr. glavonošci- lignje, hobotnice). U mehanizmu biokemijskih U regeneraciji 3. p. u kralješnjaka, očito igra važnu ulogu enzim izomeraze (vidi), koji osigurava izomerizaciju trans-retinala (ili trans-vitamina A) ponovno u cis-izomerni oblik. Međutim, konačni dokazi za postojanje takvog enzima još nisu dostupni. Sama reakcija nastajanja molekule 3.p.u prisustvu 11-cis-izomera retinala i opsina odvija se lako, bez utroška energije. Otkrivena je sposobnost izbijeljenog rodopsina da se podvrgne reakciji fosforilacije (vidi); pretpostavlja se da je ova reakcija jedna od karika u mehanizmu prilagodbe vidne stanice na svjetlost.
Bibliografija: Akkerman Yu. Biofizika, trans. s engleskog, M., 1964.; Willie K. i Det e V. Biologija, trans. s engleskog, M., 1974, bibliogr.; Konev S.V. i Volotovsky I.D. Uvod u molekularnu fotobiologiju, str. 61, Minsk, 1971.; Ostrovsky M. A. i Fedorovich I. B. Foto-inducirane promjene u fotoreceptorskoj membrani, u knjizi: Struktura i funkcije biol, membrane, ur. A. S. Troshina i sur., str. 224, M., 1975, bibliogr.; Fiziologija osjetnih sustava, ur. G.V. Gershuni, 1. dio, str. 88, L., 1971.; Biokemija i fiziologija vidnih pigmenata, ur. od H. Langera, V. a. o., 1973.; Priručnik iz senzorne fiziologije, ed. od H. A. R. Junga a. o., v. 7, t. 1-2, B., 1972.
M. A. Ostrovski.


